

An intracellular serpin regulates necrosis by inhibiting the induction and sequelae of lysosomal injury
- PMID: 17889653
- PMCID: PMC2128786
- DOI: 10.1016/j.cell.2007.07.013
An intracellular serpin regulates necrosis by inhibiting the induction and sequelae of lysosomal injury
Abstract
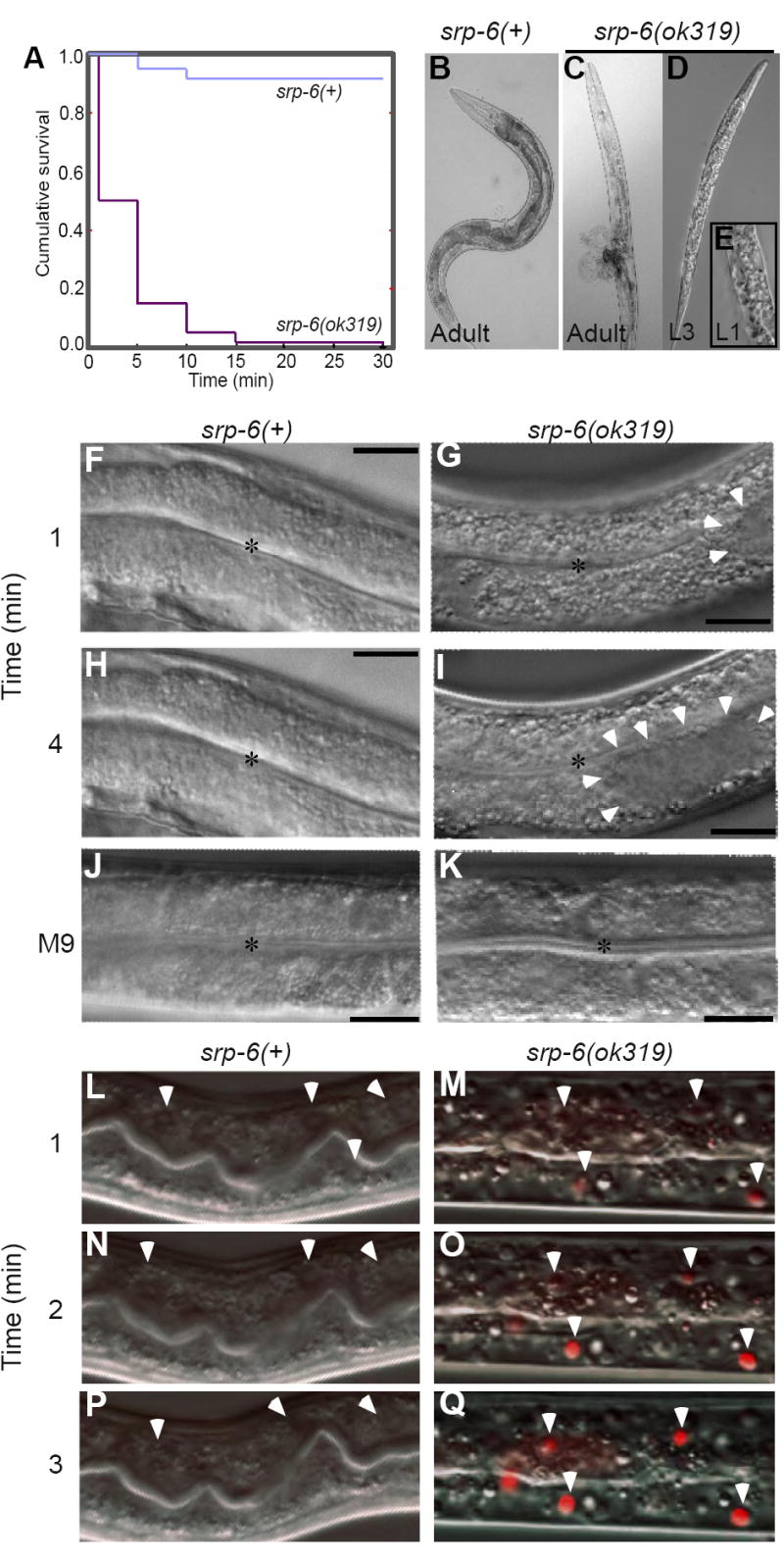
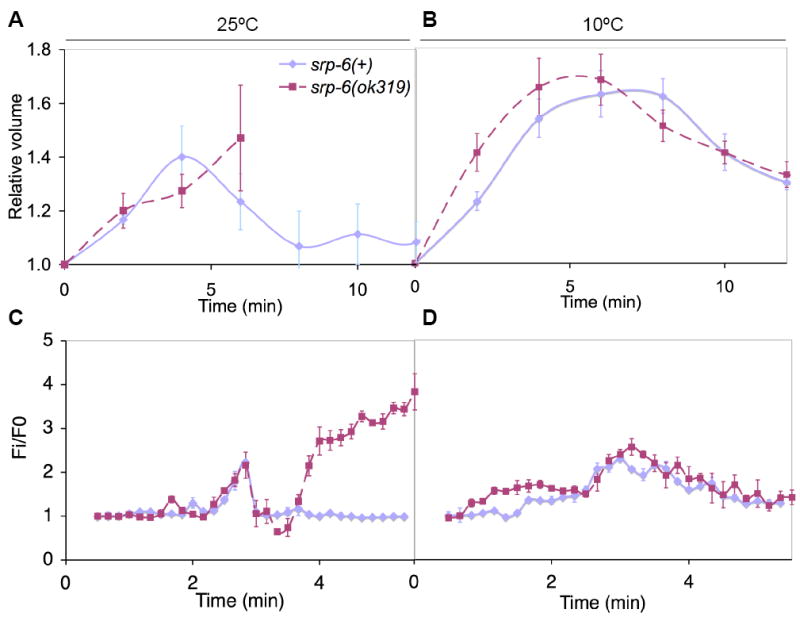
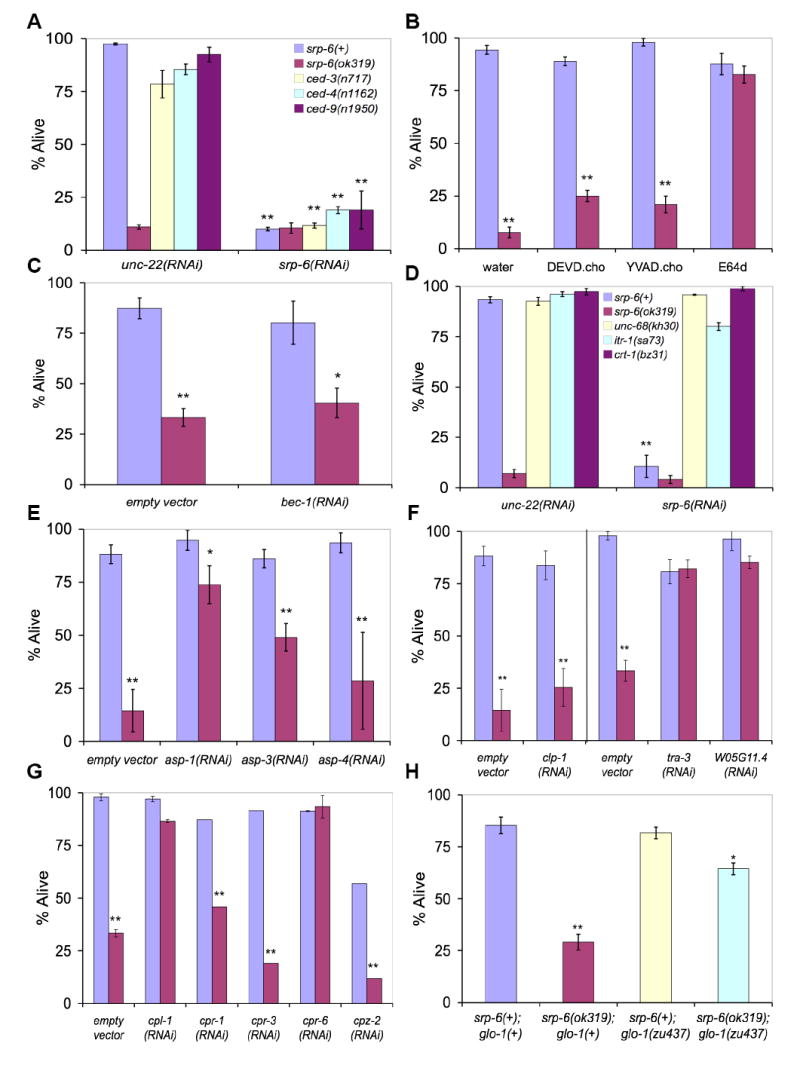
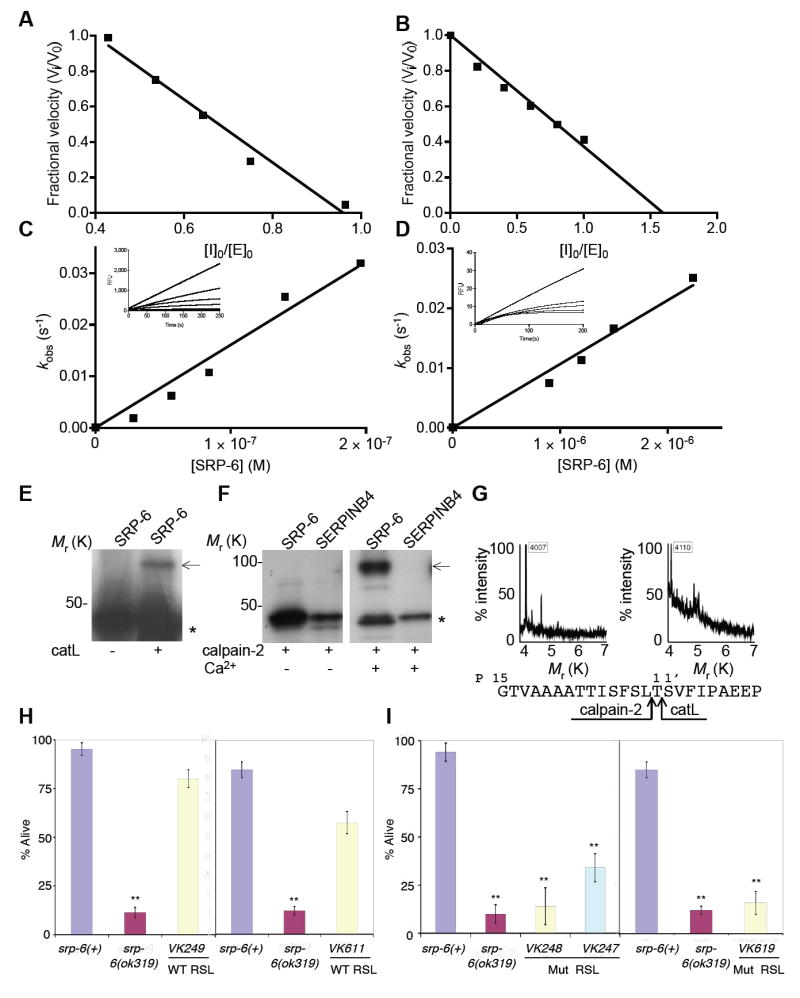
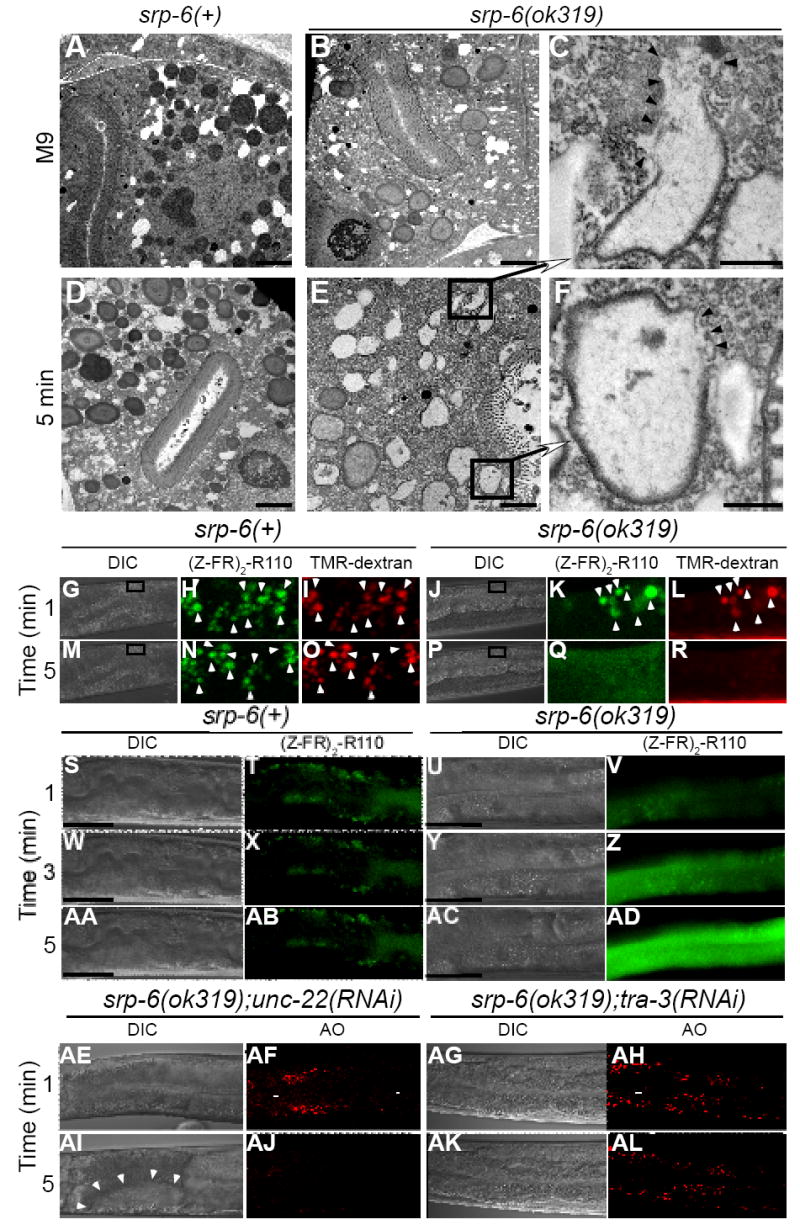
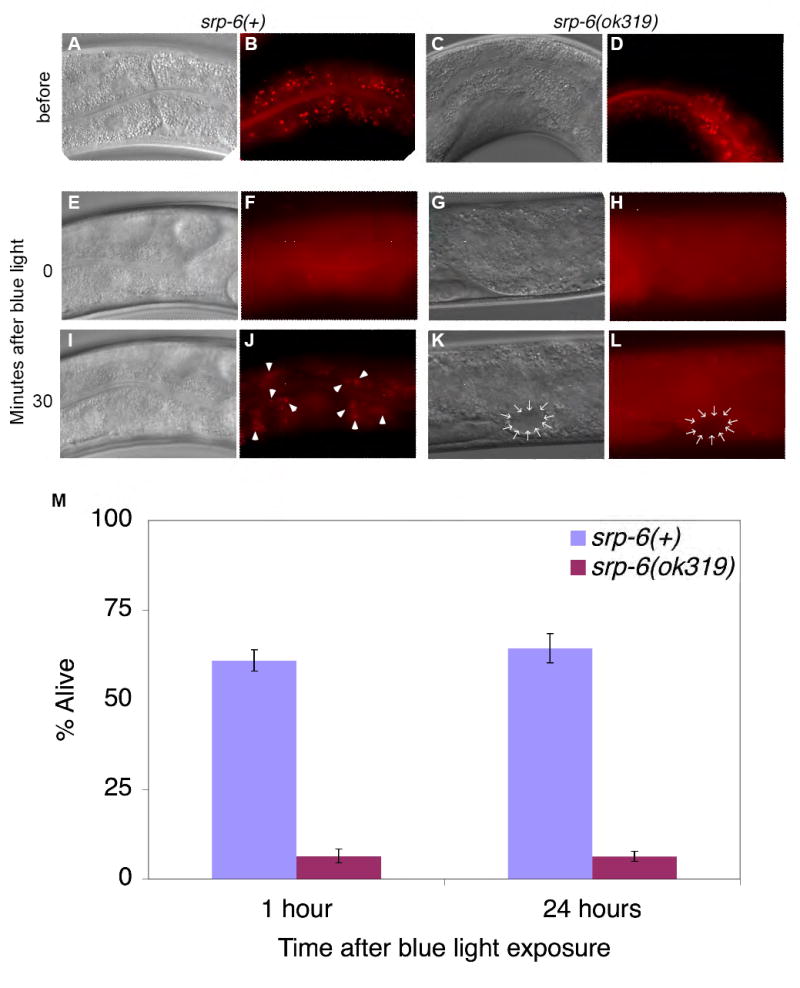
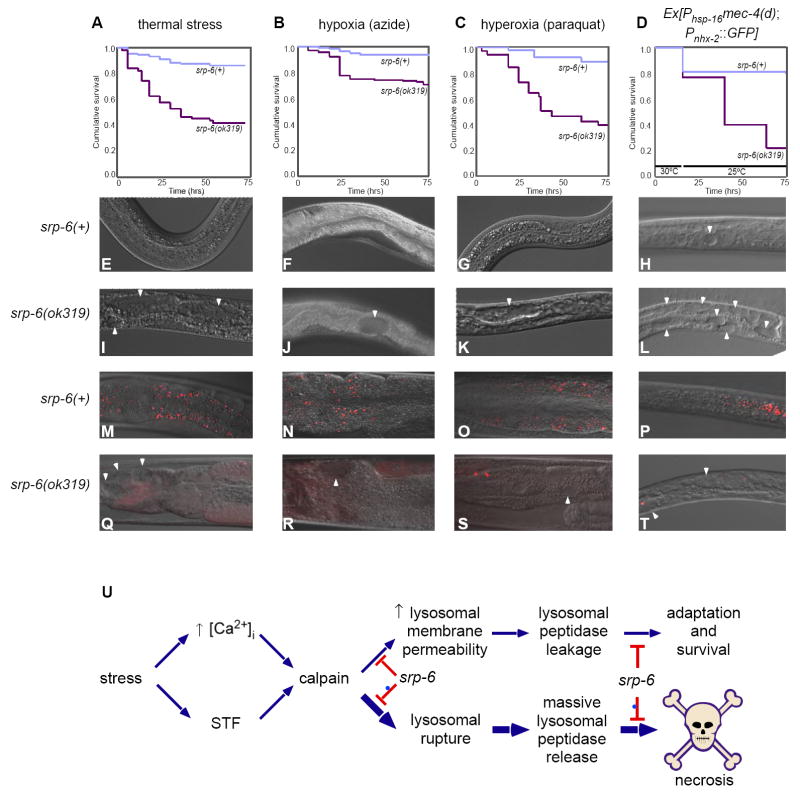
Extracellular serpins such as antithrombin and alpha1-antitrypsin are the quintessential regulators of proteolytic pathways. In contrast, the biological functions of the intracellular serpins remain obscure. We now report that the C. elegans intracellular serpin, SRP-6, exhibits a prosurvival function by blocking necrosis. Minutes after hypotonic shock, srp-6 null animals underwent a catastrophic series of events culminating in lysosomal disruption, cytoplasmic proteolysis, and death. This newly defined hypo-osmotic stress lethal (Osl) phenotype was dependent upon calpains and lysosomal cysteine peptidases, two in vitro targets of SRP-6. By protecting against both the induction of and the lethal effects from lysosomal injury, SRP-6 also blocked death induced by heat shock, oxidative stress, hypoxia, and cation channel hyperactivity. These findings suggest that multiple noxious stimuli converge upon a peptidase-driven, core stress response pathway that, in the absence of serpin regulation, triggers a lysosomal-dependent necrotic cell death routine.
Figures
References
-
- Bolger BS, Dabbas M, Lopes A, Monaghan JM. Prognostic value of preoperative squamous cell carcinoma antigen level in patients surgically treated for cervical carcinoma. Gynecol Oncol. 1997;65:309–313. - PubMed
-
- Chalfie M, Wolinsky E. The identification and suppression of inherited neurodegeneration in Caenorhabditis elegans. Nature. 1990;345:410–416. - PubMed
-
- Church FC, Pike RN, Tollefsen DM, Buckle AM, Ciaccia AV, Olson ST. Regulation of Hemostasis by Heparin-binding Serpins. In: Silverman GA, Lomas DA, editors. Molecular and Cellular Aspects of the Serpinopathies and Disorders of Serpin Activity. Singapore: World Scientific Publishing Co. Pte. Ltd.; 2007. pp. 509–554.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
