

Sir2 deacetylates histone H3 lysine 56 to regulate telomeric heterochromatin structure in yeast
- PMID: 17889663
- PMCID: PMC2048486
- DOI: 10.1016/j.molcel.2007.07.021
Sir2 deacetylates histone H3 lysine 56 to regulate telomeric heterochromatin structure in yeast
Abstract
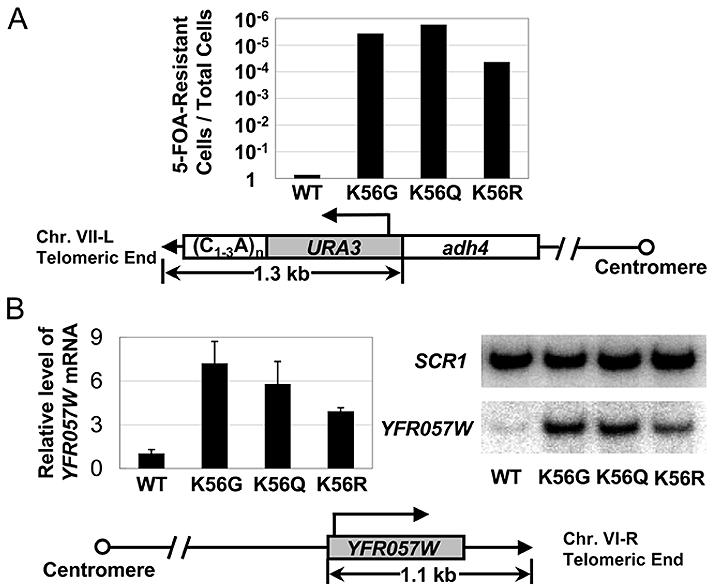
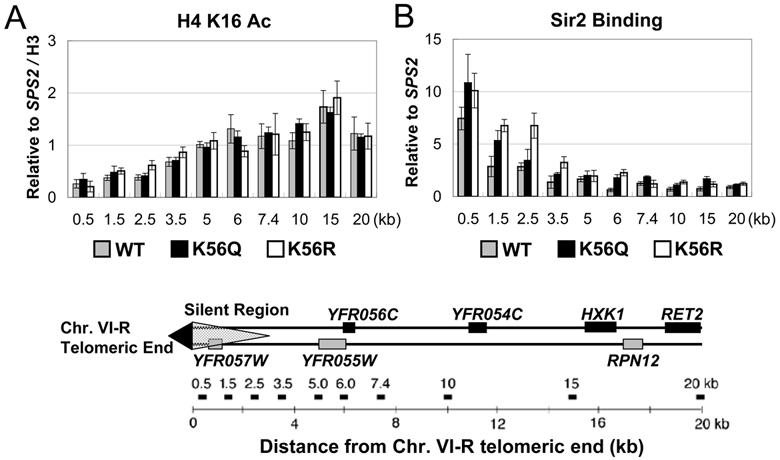
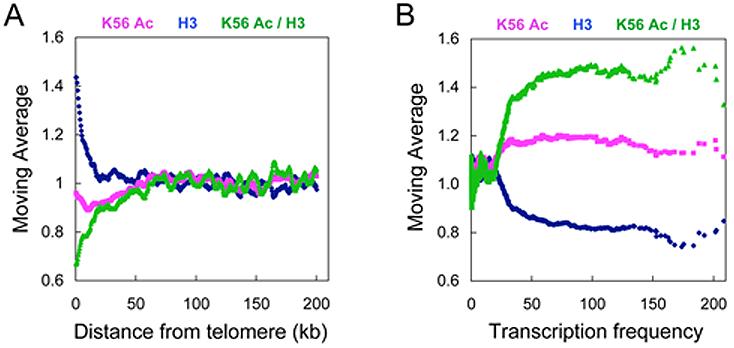
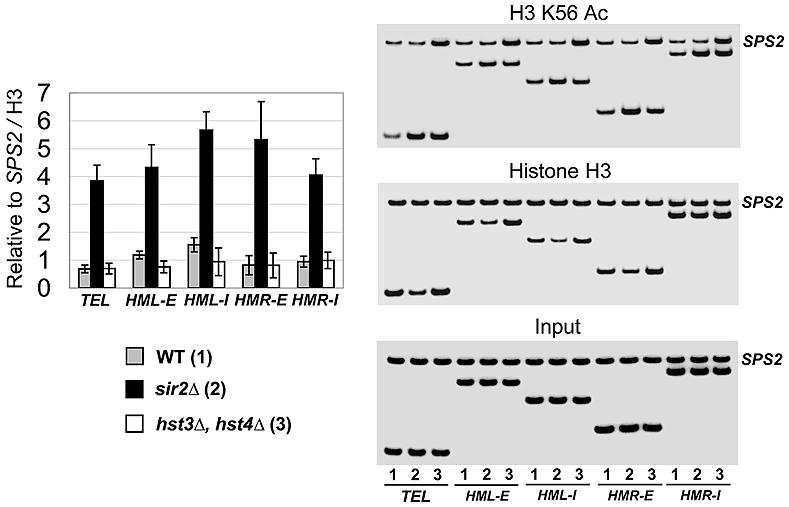
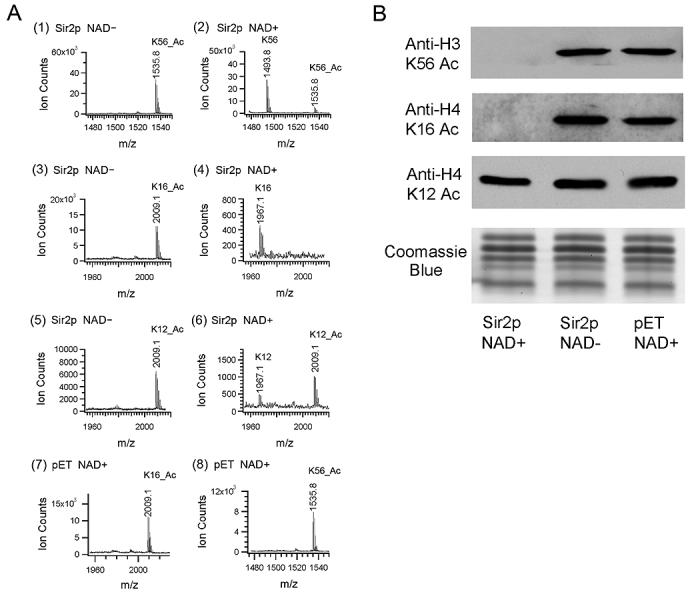
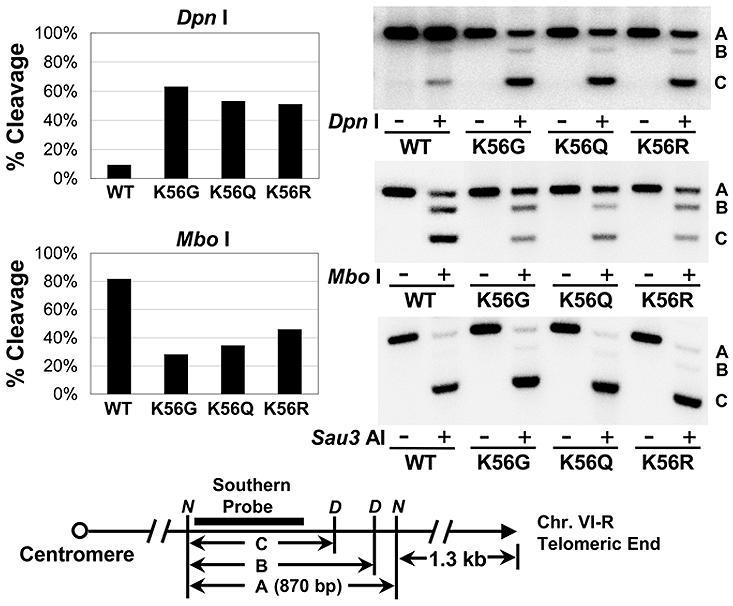
At telomeric heterochromatin in yeast, the Sir protein complex spreads from Rap1 sites to silence adjacent genes. This cascade is believed to occur when Sir2, an NAD(+)-dependent enzyme, deacetylates histone H3 and H4 N termini, in particular histone H4 K16, enabling more Sir protein binding. Lysine 56 of histone H3 is located at the entry-exit points of the DNA superhelix surrounding the nucleosome, where it may control DNA compaction. We have found that K56 substitutions disrupt silencing severely without decreasing Sir protein binding at the telomere. Our in vitro and in vivo data indicate that Sir2 deacetylates K56 directly in telomeric heterochromatin to compact chromatin and prevent access to RNA polymerase and ectopic bacterial dam methylase. Since the spread of Sir proteins is necessary but not sufficient for silencing, we propose that silencing occurs when Sir2 deacetylates H3 K56 to close the nucleosomal entry-exit gates, enabling compaction of heterochromatin.
Figures
Similar articles
-
Rap1-Sir4 binding independent of other Sir, yKu, or histone interactions initiates the assembly of telomeric heterochromatin in yeast.Genes Dev. 2002 Jun 15;16(12):1528-39. doi: 10.1101/gad.988802. Genes Dev. 2002. PMID: 12080091 Free PMC article.
-
Reconstitution of heterochromatin-dependent transcriptional gene silencing.Mol Cell. 2009 Sep 24;35(6):769-81. doi: 10.1016/j.molcel.2009.07.030. Mol Cell. 2009. PMID: 19782027 Free PMC article.
-
The C-terminus of histone H2B is involved in chromatin compaction specifically at telomeres, independently of its monoubiquitylation at lysine 123.PLoS One. 2011;6(7):e22209. doi: 10.1371/journal.pone.0022209. Epub 2011 Jul 29. PLoS One. 2011. PMID: 21829450 Free PMC article.
-
A model for step-wise assembly of heterochromatin in yeast.Novartis Found Symp. 2004;259:48-56; discussion 56-62, 163-9. Novartis Found Symp. 2004. PMID: 15171246 Review.
-
Molecular model for telomeric heterochromatin in yeast.Curr Opin Cell Biol. 1997 Jun;9(3):383-7. doi: 10.1016/s0955-0674(97)80011-7. Curr Opin Cell Biol. 1997. PMID: 9159071 Review.
Cited by
-
H4K16 acetylation affects recombination and ncRNA transcription at rDNA in Saccharomyces cerevisiae.Mol Biol Cell. 2012 Jul;23(14):2770-81. doi: 10.1091/mbc.E12-02-0095. Epub 2012 May 23. Mol Biol Cell. 2012. PMID: 22621897 Free PMC article.
-
Acetylated histone H3K56 interacts with Oct4 to promote mouse embryonic stem cell pluripotency.Proc Natl Acad Sci U S A. 2013 Jul 9;110(28):11493-8. doi: 10.1073/pnas.1309914110. Epub 2013 Jun 24. Proc Natl Acad Sci U S A. 2013. PMID: 23798425 Free PMC article.
-
Replisome function during replicative stress is modulated by histone h3 lysine 56 acetylation through Ctf4.Genetics. 2015 Apr;199(4):1047-63. doi: 10.1534/genetics.114.173856. Epub 2015 Feb 18. Genetics. 2015. PMID: 25697176 Free PMC article.
-
Discovery of histone modification crosstalk networks by stable isotope labeling of amino acids in cell culture mass spectrometry (SILAC MS).Mol Cell Proteomics. 2013 Aug;12(8):2048-59. doi: 10.1074/mcp.M112.026716. Epub 2013 Apr 15. Mol Cell Proteomics. 2013. PMID: 23592332 Free PMC article.
-
Histone H3 N-terminus regulates higher order structure of yeast heterochromatin.Proc Natl Acad Sci U S A. 2009 Aug 11;106(32):13153-9. doi: 10.1073/pnas.0906866106. Epub 2009 Aug 3. Proc Natl Acad Sci U S A. 2009. PMID: 19666585 Free PMC article.
References
-
- Allshire RC, Javerzat JP, Redhead NJ, Cranston G. Position effect variegation at fission yeast centromeres. Cell. 1994;76:157–169. - PubMed
-
- Aparicio OM, Billington BL, Gottschling DE. Modifiers of position effect are shared between telomeric and silent mating-type loci in S. cerevisiae. Cell. 1991;66:1279–1287. - PubMed
-
- Boeger H, Griesenbeck J, Strattan JS, Kornberg RD. Nucleosomes unfold completely at a transcriptionally active promoter. Mol Cell. 2003;11:1587–1598. - PubMed
-
- Braunstein M, Rose AB, Holmes SG, Allis CD, Broach JR. Transcriptional silencing in yeast is associated with reduced nucleosome acetylation. Genes Dev. 1993;7:592–604. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
