

Neural specializations for speech and pitch: moving beyond the dichotomies
- PMID: 17890188
- PMCID: PMC2606798
- DOI: 10.1098/rstb.2007.2161
Neural specializations for speech and pitch: moving beyond the dichotomies
Abstract
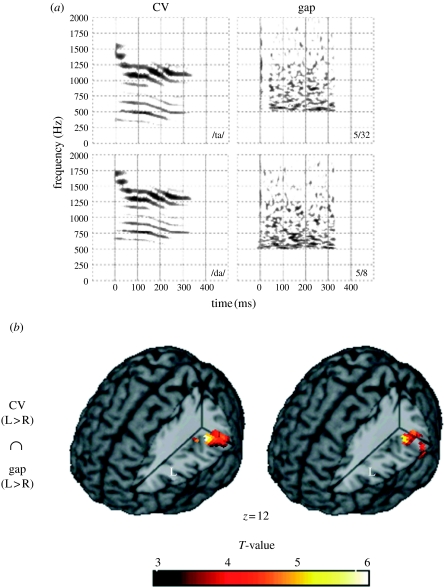
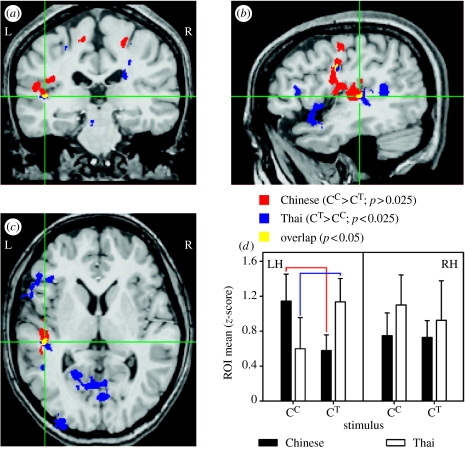
The idea that speech processing relies on unique, encapsulated, domain-specific mechanisms has been around for some time. Another well-known idea, often espoused as being in opposition to the first proposal, is that processing of speech sounds entails general-purpose neural mechanisms sensitive to the acoustic features that are present in speech. Here, we suggest that these dichotomous views need not be mutually exclusive. Specifically, there is now extensive evidence that spectral and temporal acoustical properties predict the relative specialization of right and left auditory cortices, and that this is a parsimonious way to account not only for the processing of speech sounds, but also for non-speech sounds such as musical tones. We also point out that there is equally compelling evidence that neural responses elicited by speech sounds can differ depending on more abstract, linguistically relevant properties of a stimulus (such as whether it forms part of one's language or not). Tonal languages provide a particularly valuable window to understand the interplay between these processes. The key to reconciling these phenomena probably lies in understanding the interactions between afferent pathways that carry stimulus information, with top-down processing mechanisms that modulate these processes. Although we are still far from the point of having a complete picture, we argue that moving forward will require us to abandon the dichotomy argument in favour of a more integrated approach.
Figures





References
-
- Bavelier D, Corina D.P, Neville H.J. Brain and language: a perspective from sign language. Neuron. 1998;21:275–278. doi:10.1016/S0896-6273(00)80536-X - DOI - PubMed
-
- Belin P, Zilbovicius M, Crozier S, Thivard L, Fontaine A, Masure M.-C, Samson Y. Lateralization of speech and auditory temporal processing. J. Cogn. Neurosci. 1998;10:536–540. doi:10.1162/089892998562834 - DOI - PubMed
-
- Binder J, Frost J, Hammeke T, Bellgowan P, Springer J, Kaufman J, Possing J. Human temporal lobe activation by speech and nonspeech sounds. Cereb. Cortex. 2000;10:512–528. doi:10.1093/cercor/10.5.512 - DOI - PubMed
-
- Blumstein S.E. The neurobiology of the sound structure of language. In: Gazzaniga M.S, editor. The cognitive neurosciences. MIT Press; Cambridge, MA: 1994. pp. 915–929.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources