

How moist heat kills spores of Bacillus subtilis
- PMID: 17890306
- PMCID: PMC2168948
- DOI: 10.1128/JB.01242-07
How moist heat kills spores of Bacillus subtilis
Abstract
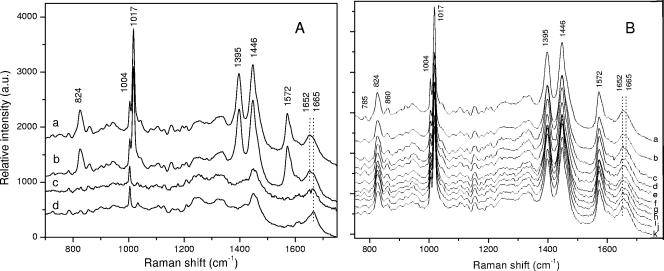
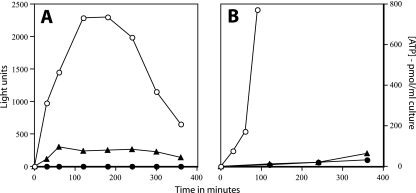
Populations of Bacillus subtilis spores in which 90 to 99.9% of the spores had been killed by moist heat gave only two fractions on equilibrium density gradient centrifugation: a fraction comprised of less dense spores that had lost their dipicolinic acid (DPA), undergone significant protein denaturation, and were all dead and a fraction with the same higher density as that of unheated spores. The latter fraction from heat-killed spore populations retained all of its DPA, but >/=98% of the spores could be dead. The dead spores that retained DPA germinated relatively normally with nutrient and nonnutrient germinants, but the outgrowth of these germinated spores was significantly compromised, perhaps because they had suffered damage to some proteins such that metabolic activity during outgrowth was greatly decreased. These results indicate that DPA release takes place well after spore killing by moist heat and that DPA release during moist-heat treatment is an all-or-nothing phenomenon; these findings also suggest that damage to one or more key spore proteins causes spore killing by moist heat.
Figures





References
-
- Atrih, A., and S. J. Foster. 2001. In vivo roles of the germination-specific lytic enzymes of Bacillus subtilis 168. Microbiology 147:2925-2932. - PubMed
-
- Cazemier, A. E., S. F. Wagenaars, and P. F. ter Steeg. 2001. Effect of sporulation and recovery medium on the heat resistance and amount of injury of spores from spoilage bacilli. J. Appl. Microbiol. 90:761-770. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
