

Molecular evolution of the vertebrate mechanosensory cell and ear
- PMID: 17891725
- PMCID: PMC3918877
- DOI: 10.1387/ijdb.072367bf
Molecular evolution of the vertebrate mechanosensory cell and ear
Abstract
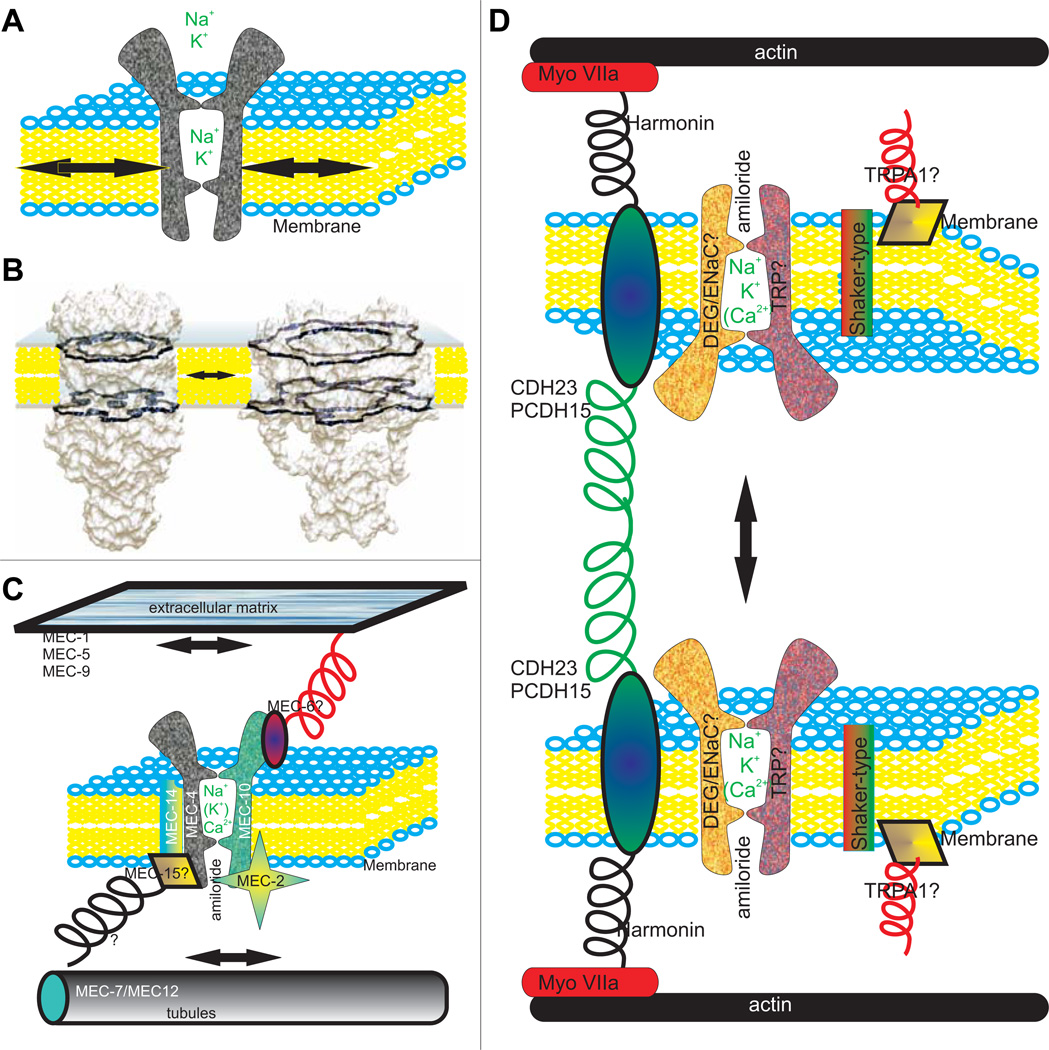
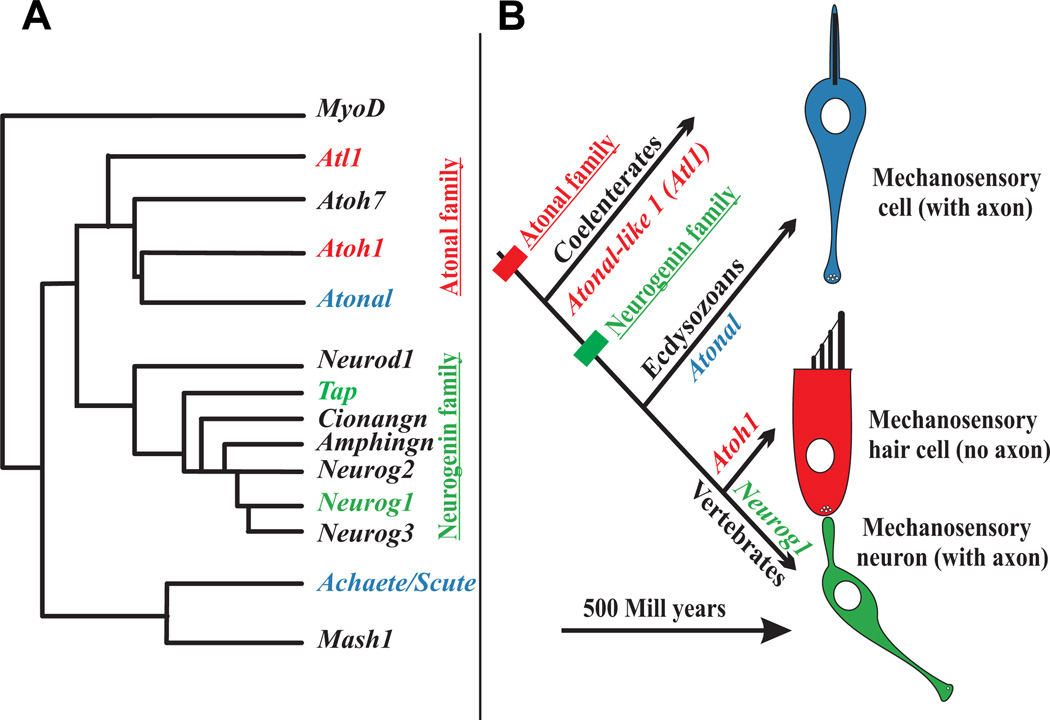
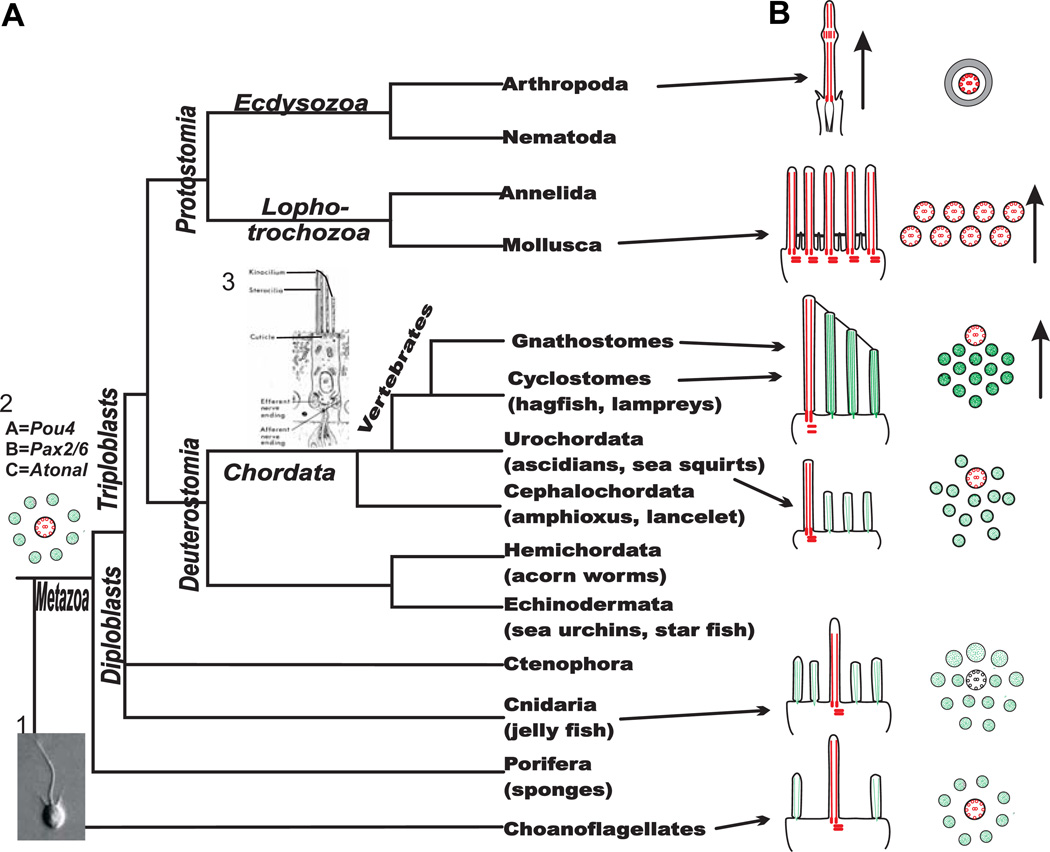
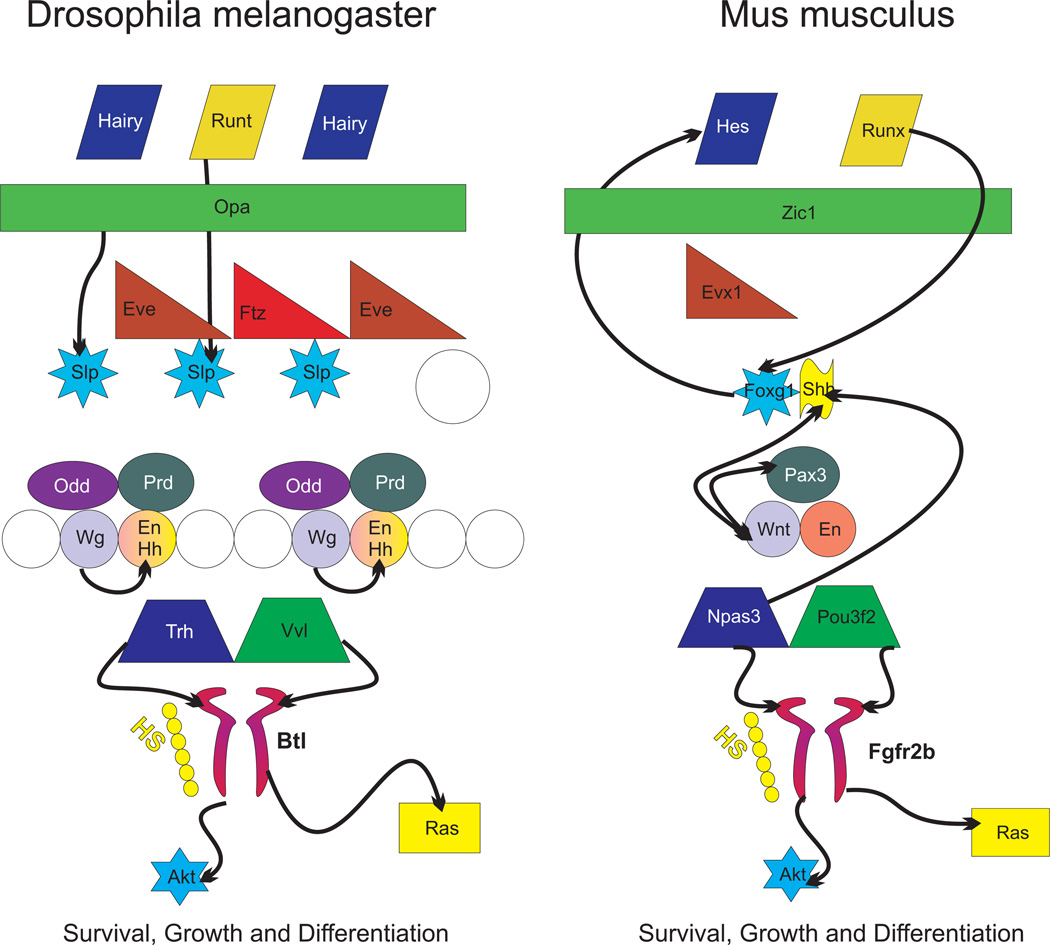
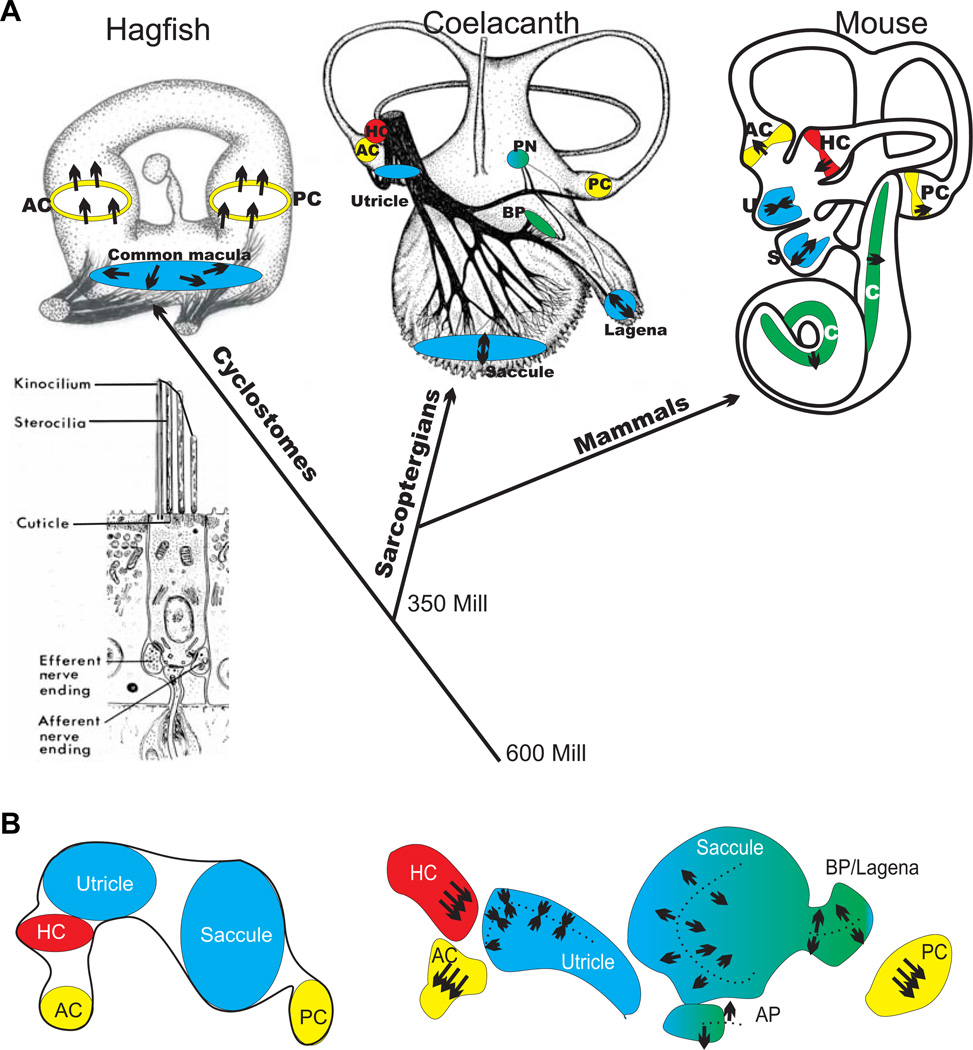
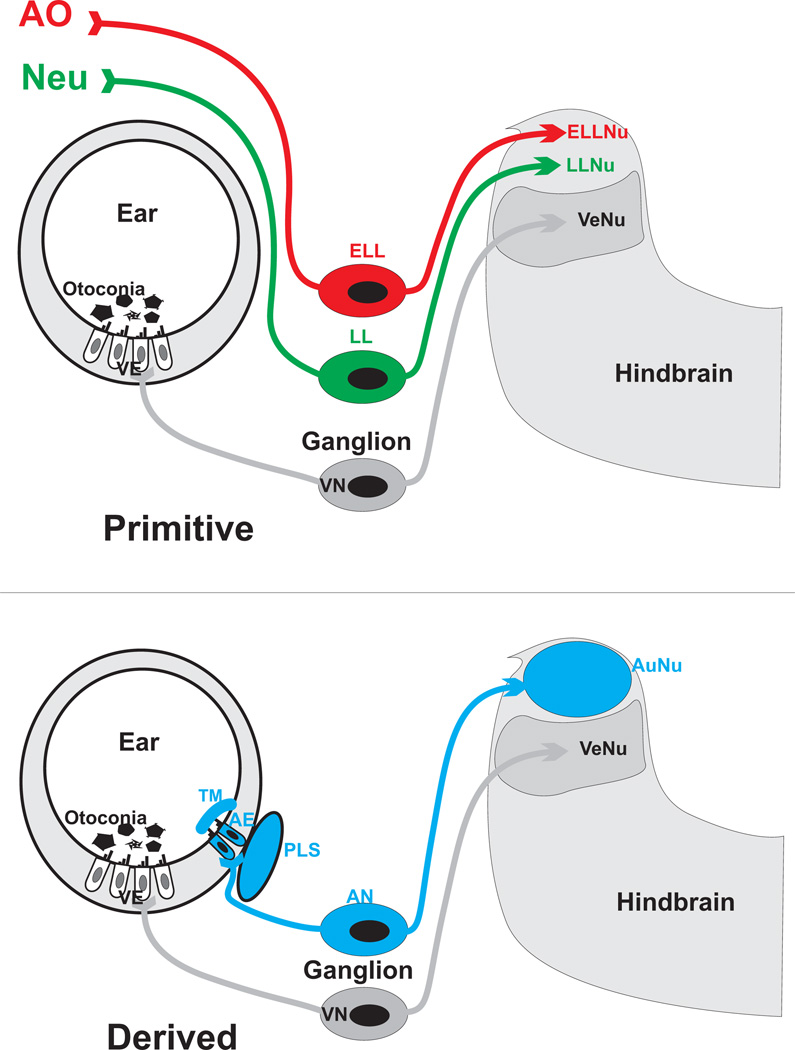
The molecular basis of mechanosensation, mechanosensory cell development and mechanosensory organ development is reviewed with an emphasis on its evolution. In contrast to eye evolution and development, which apparently modified a genetic program through intercalation of genes between the master control genes on the top (Pax6, Eya1, Six1) of the hierarchy and the structural genes (rhodopsin) at the bottom, the as yet molecularly unknown mechanosensory channel precludes such a firm conclusion for mechanosensors. However, recent years have seen the identification of several structural genes which are involved in mechanosensory tethering and several transcription factors controlling mechanosensory cell and organ development; these warrant the interpretation of available data in very much the same fashion as for eye evolution: molecular homology combined with potential morphological parallelism. This assertion of molecular homology is strongly supported by recent findings of a highly conserved set of microRNAs that appear to be associated with mechanosensory cell development across phyla. The conservation of transcription factors and their regulators fits very well to the known or presumed mechanosensory specializations which can be mostly grouped as variations of a common cellular theme. Given the widespread distribution of the molecular ability to form mechanosensory cells, it comes as no surprise that structurally different mechanosensory organs evolved in different phyla, presenting a variation of a common theme specified by a conserved set of transcription factors in their cellular development. Within vertebrates and arthropods, some mechanosensory organs evolved into auditory organs, greatly increasing sensitivity to sound through modifications of accessory structures to direct sound to the specific sensory epithelia. However, while great attention has been paid to the evolution of these accessory structures in vertebrate fossils, comparatively less attention has been spent on the evolution of the inner ear and the central auditory system. Recent advances in our molecular understanding of ear and brain development provide novel avenues to this neglected aspect of auditory neurosensory evolution.
Figures
References
-
- Alonso CR, Wilkins AS. The molecular elements that underlie developmental evolution. Nat Rev Genet. 2005;6:709–715. - PubMed
-
- Alvarez Y, Alonso MT, Vendrell V, Zelarayan LC, Chamero P, Theil T, Bosl MR, Kato S, Maconochie M, Riethmacher D, et al. Requirements for FGF3 and FGF10 during inner ear formation. Development. 2003;130:6329–6338. - PubMed
-
- Arendt D, Tessmar-Raible K, Snyman H, Dorresteijn AW, Wittbrodt J. Ciliary photoreceptors with a vertebrate-type opsin in an invertebrate brain. Science. 2004;306:869–871. - PubMed
-
- Arkett SA, Mackie GO, Meech RW. Hair cell mechanoreception in a jellyfish Aglantha digitale. Journal of Experimental Biology. 1988;135:329–342.
-
- Bailey AP, Bhattacharyya S, Bronner-Fraser M, Streit A. Lens specification is the ground state of all sensory placodes, from which FGF promotes olfactory identity. Dev Cell. 2006;11:505–517. - PubMed
