

Accessibility of targeted DHPR sites to streptavidin and functional effects of binding on EC coupling
- PMID: 17893191
- PMCID: PMC2151652
- DOI: 10.1085/jgp.200609730
Accessibility of targeted DHPR sites to streptavidin and functional effects of binding on EC coupling
Abstract
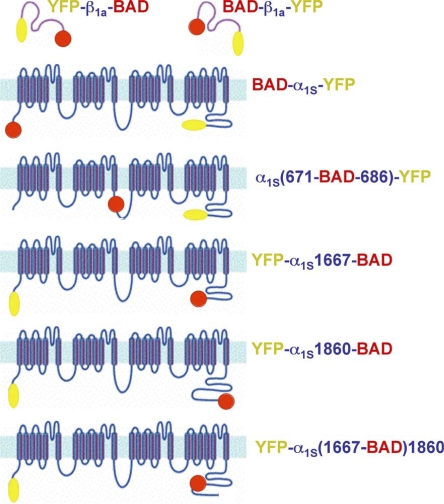
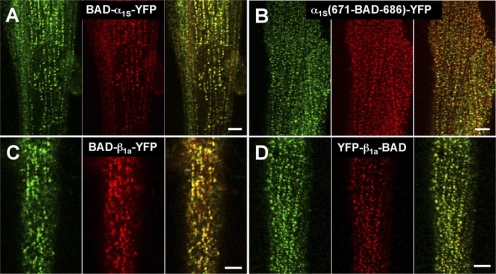
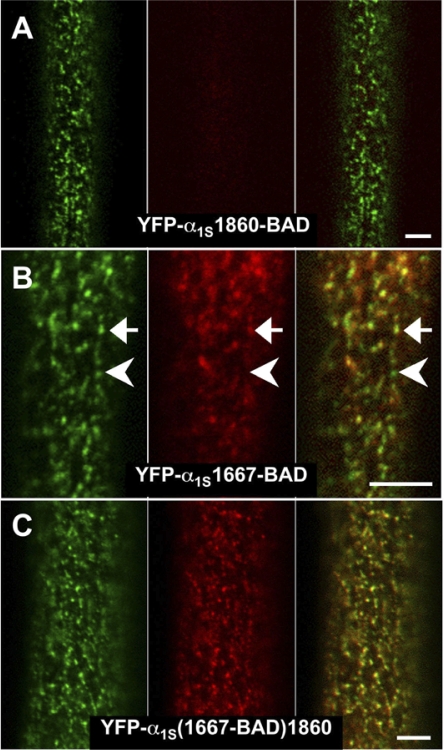
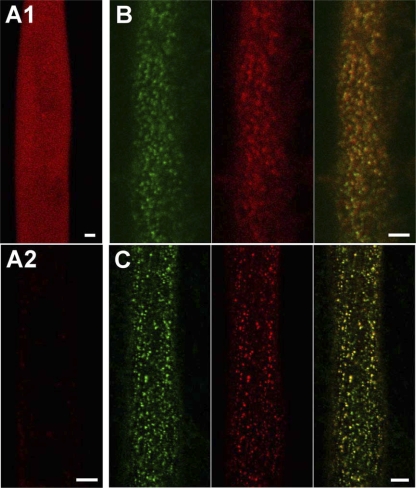
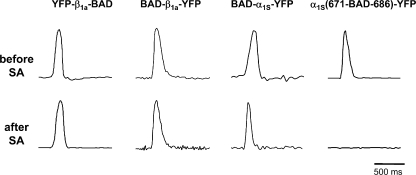
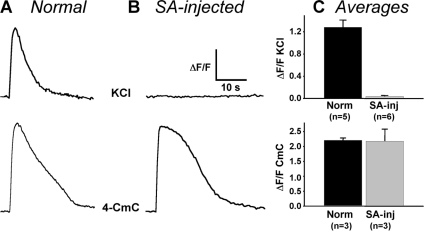
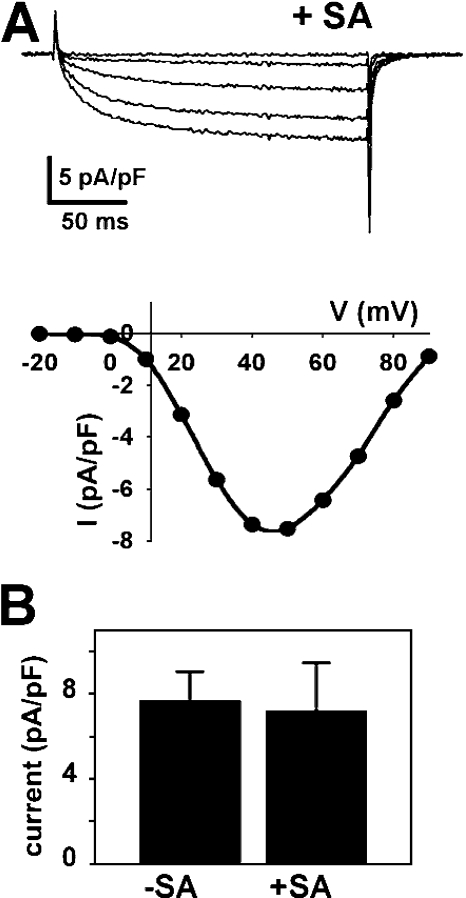
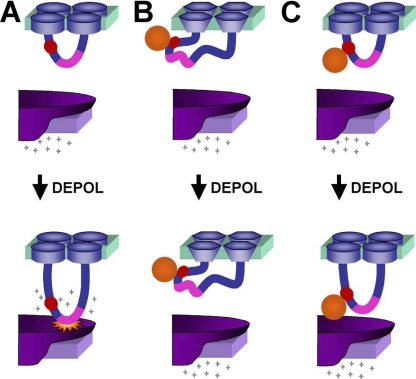
In skeletal muscle, the dihydropyridine receptor (DHPR) in the plasma membrane (PM) serves as a Ca(2+) channel and as the voltage sensor for excitation-contraction (EC coupling), triggering Ca(2+) release via the type 1 ryanodine receptor (RyR1) in the sarcoplasmic reticulum (SR) membrane. In addition to being functionally linked, these two proteins are also structurally linked to one another, but the identity of these links remains unknown. As an approach to address this issue, we have expressed DHPR alpha(1S) or beta(1a) subunits, with a biotin acceptor domain fused to targeted sites, in myotubes null for the corresponding, endogenous DHPR subunit. After saponin permeabilization, the approximately 60-kD streptavidin molecule had access to the beta(1a) N and C termini and to the alpha(1S) N terminus and proximal II-III loop (residues 671-686). Steptavidin also had access to these sites after injection into living myotubes. However, sites of the alpha(1S) C terminus were either inaccessible or conditionally accessible in saponin- permeabilized myotubes, suggesting that these C-terminal regions may exist in conformations that are occluded by other proteins in PM/SR junction (e.g., RyR1). The binding of injected streptavidin to the beta(1a) N or C terminus, or to the alpha(1S) N terminus, had no effect on electrically evoked contractions. By contrast, binding of streptavidin to the proximal alpha(1S) II-III loop abolished such contractions, without affecting agonist-induced Ca(2+) release via RyR1. Moreover, the block of EC coupling did not appear to result from global distortion of the DHPR and supports the hypothesis that conformational changes of the alpha(1S) II-III loop are necessary for EC coupling in skeletal muscle.
Figures
Similar articles
-
Fluorescence resonance energy transfer (FRET) indicates that association with the type I ryanodine receptor (RyR1) causes reorientation of multiple cytoplasmic domains of the dihydropyridine receptor (DHPR) α(1S) subunit.J Biol Chem. 2012 Nov 30;287(49):41560-8. doi: 10.1074/jbc.M112.404194. Epub 2012 Oct 15. J Biol Chem. 2012. PMID: 23071115 Free PMC article.
-
Ca2+-dependent excitation-contraction coupling triggered by the heterologous cardiac/brain DHPR beta2a-subunit in skeletal myotubes.Biophys J. 2003 Dec;85(6):3739-57. doi: 10.1016/S0006-3495(03)74790-0. Biophys J. 2003. PMID: 14645065 Free PMC article.
-
Rem uncouples excitation-contraction coupling in adult skeletal muscle fibers.J Gen Physiol. 2015 Jul;146(1):97-108. doi: 10.1085/jgp.201411314. Epub 2015 Jun 15. J Gen Physiol. 2015. PMID: 26078055 Free PMC article.
-
Bridging the myoplasmic gap: recent developments in skeletal muscle excitation-contraction coupling.J Muscle Res Cell Motil. 2007;28(4-5):275-83. doi: 10.1007/s10974-007-9118-5. Epub 2007 Sep 26. J Muscle Res Cell Motil. 2007. PMID: 17899404 Review.
-
Functional equivalence of dihydropyridine receptor alpha1S and beta1a subunits in triggering excitation-contraction coupling in skeletal muscle.Biol Res. 2004;37(4):565-75. doi: 10.4067/s0716-97602004000400010. Biol Res. 2004. PMID: 15709683 Review.
Cited by
-
Fluorescence resonance energy transfer (FRET) indicates that association with the type I ryanodine receptor (RyR1) causes reorientation of multiple cytoplasmic domains of the dihydropyridine receptor (DHPR) α(1S) subunit.J Biol Chem. 2012 Nov 30;287(49):41560-8. doi: 10.1074/jbc.M112.404194. Epub 2012 Oct 15. J Biol Chem. 2012. PMID: 23071115 Free PMC article.
-
Ca(V)1.1: The atypical prototypical voltage-gated Ca²⁺ channel.Biochim Biophys Acta. 2013 Jul;1828(7):1587-97. doi: 10.1016/j.bbamem.2012.09.007. Epub 2012 Sep 13. Biochim Biophys Acta. 2013. PMID: 22982493 Free PMC article. Review.
-
Reciprocal dihydropyridine and ryanodine receptor interactions in skeletal muscle activation.J Muscle Res Cell Motil. 2011 Nov;32(3):171-202. doi: 10.1007/s10974-011-9262-9. Epub 2011 Oct 13. J Muscle Res Cell Motil. 2011. PMID: 21993921 Review.
-
Bimolecular fluorescence complementation and targeted biotinylation provide insight into the topology of the skeletal muscle Ca ( 2+) channel β1a subunit.Channels (Austin). 2012 Jan-Feb;6(1):26-40. doi: 10.4161/chan.18916. Epub 2012 Jan 1. Channels (Austin). 2012. PMID: 22522946 Free PMC article.
-
Looking for answers to EC coupling's persistent questions.J Gen Physiol. 2010 Jul;136(1):7-12. doi: 10.1085/jgp.201010461. J Gen Physiol. 2010. PMID: 20584887 Free PMC article. No abstract available.
References
-
- Bannister, R.A., and K.G. Beam. 2005. The α1S N-terminus is not essential for bi-directional coupling with RyR1. Biochem. Biophys. Res. Commun. 336:134–141. - PubMed
-
- Beam, K.G., and C. Franzini-Armstrong. 1997. Functional and structural approaches to the study of excitation-contraction coupling. Methods Cell Biol. 52:283–306. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
