

Superior control of HIV-1 replication by CD8+ T cells is reflected by their avidity, polyfunctionality, and clonal turnover
- PMID: 17893201
- PMCID: PMC2118466
- DOI: 10.1084/jem.20070784
Superior control of HIV-1 replication by CD8+ T cells is reflected by their avidity, polyfunctionality, and clonal turnover
Abstract
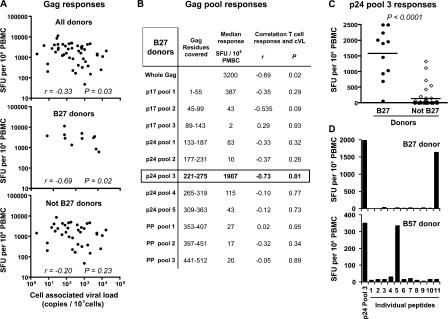
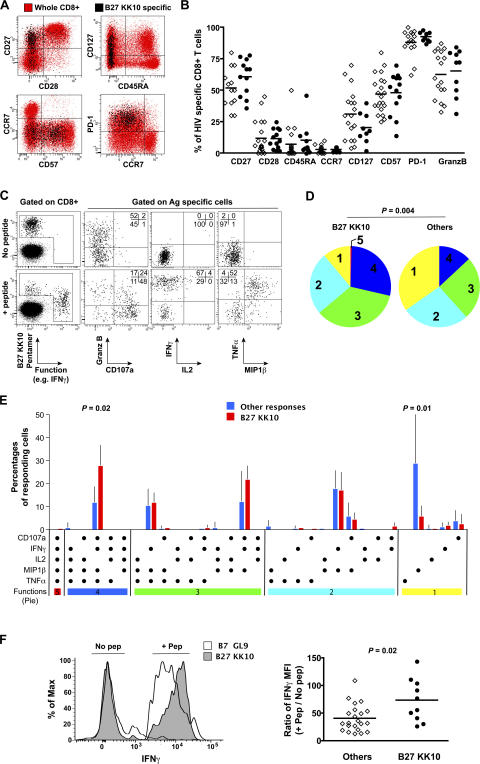
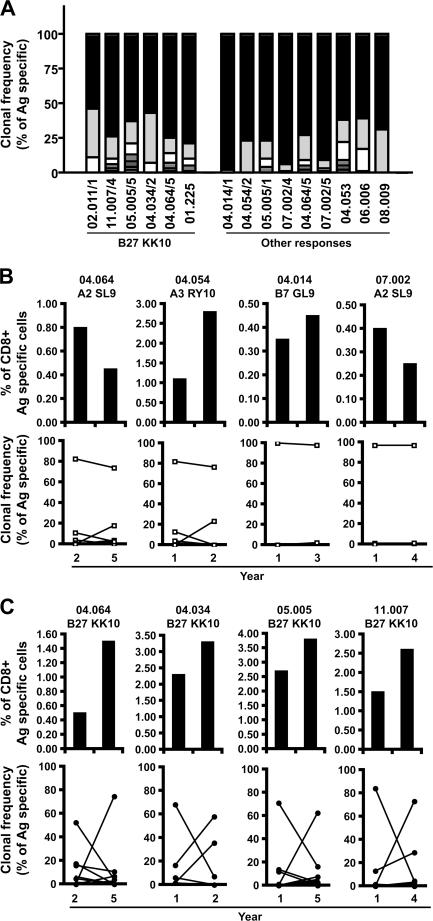
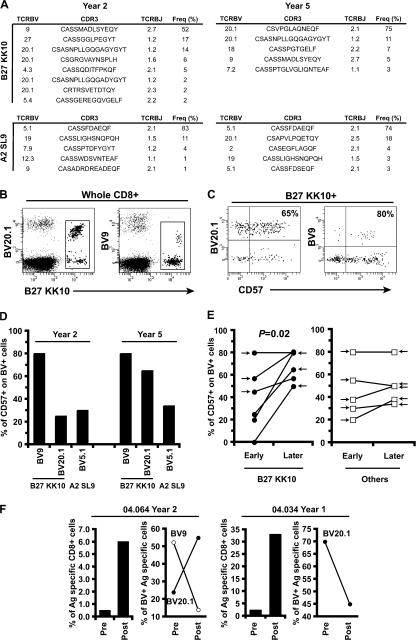
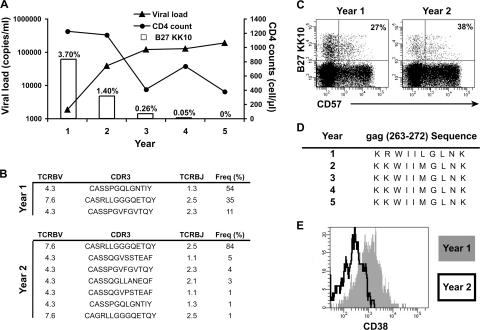
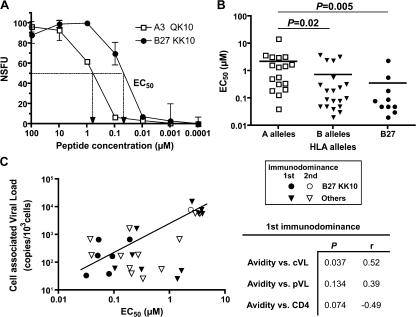
The key attributes of CD8+ T cell protective immunity in human immunodeficiency virus (HIV) infection remain unclear. We report that CD8+ T cell responses specific for Gag and, in particular, the immunodominant p24 epitope KK10 correlate with control of HIV-1 replication in human histocompatibility leukocyte antigen (HLA)-B27 patients. To understand further the nature of CD8+ T cell-mediated antiviral efficacy, we performed a comprehensive study of CD8+ T cells specific for the HLA-B27-restricted epitope KK10 in chronic HIV-1 infection based on the use of multiparametric flow cytometry together with molecular clonotypic analysis and viral sequencing. We show that B27-KK10-specific CD8+ T cells are characterized by polyfunctional capabilities, increased clonal turnover, and superior functional avidity. Such attributes are interlinked and constitute the basis for effective control of HIV-1 replication. These data on the features of effective CD8+ T cells in HIV infection may aid in the development of successful T cell vaccines.
Figures
References
-
- Gea-Banacloche, J.C., S.A. Migueles, L. Martino, W.L. Shupert, A.C. McNeil, M.S. Sabbaghian, L. Ehler, C. Prussin, R. Stevens, L. Lambert, et al. 2000. Maintenance of large numbers of virus-specific CD8+ T cells in HIV-infected progressors and long-term nonprogressors. J. Immunol. 165:1082–1092. - PubMed
-
- Betts, M.R., D.R. Ambrozak, D.C. Douek, S. Bonhoeffer, J.M. Brenchley, J.P. Casazza, R.A. Koup, and L.J. Picker. 2001. Analysis of total human immunodeficiency virus (HIV)-specific CD4(+) and CD8(+) T-cell responses: relationship to viral load in untreated HIV infection. J. Virol. 75:11983–11991. - PMC - PubMed
-
- Kaslow, R.A., M. Carrington, R. Apple, L. Park, A. Munoz, A.J. Saah, J.J. Goedert, C. Winkler, S.J. O'Brien, C. Rinaldo, et al. 1996. Influence of combinations of human major histocompatibility complex genes on the course of HIV-1 infection. Nat. Med. 2:405–411. - PubMed
-
- Magierowska, M., I. Theodorou, P. Debre, F. Sanson, B. Autran, Y. Riviere, D. Charron, and D. Costagliola. 1999. Combined genotypes of CCR5, CCR2, SDF1, and HLA genes can predict the long-term nonprogressor status in human immunodeficiency virus-1-infected individuals. Blood. 93:936–941. - PubMed
-
- O'Brien, S.J., G.W. Nelson, C.A. Winkler, and M.W. Smith. 2000. Polygenic and multifactorial disease gene association in man: Lessons from AIDS. Annu. Rev. Genet. 34:563–591. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
