

APC mutations lead to cytokinetic failures in vitro and tetraploid genotypes in Min mice
- PMID: 17893240
- PMCID: PMC2064647
- DOI: 10.1083/jcb.200703186
APC mutations lead to cytokinetic failures in vitro and tetraploid genotypes in Min mice
Abstract
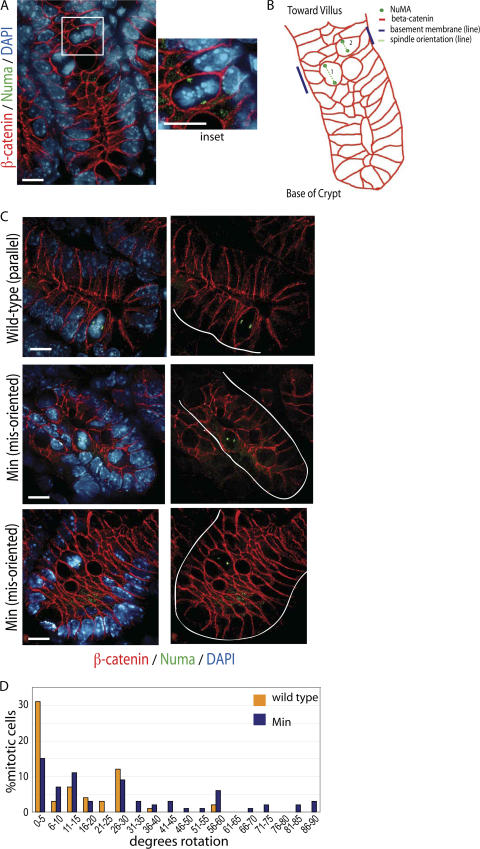
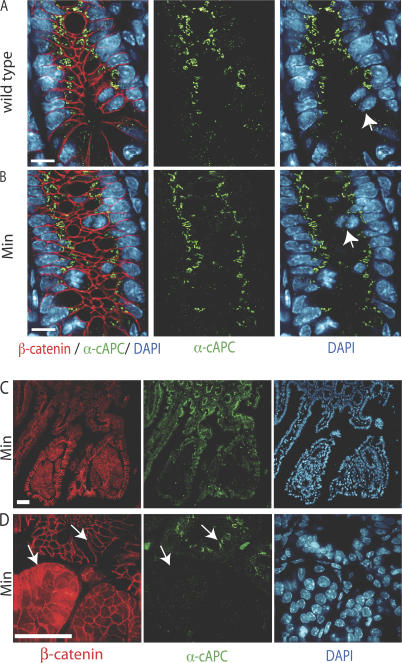
Previous research has proposed that genomic instability contributes to cancer progression, with its initiation linked to tetraploid cell formation (Duesberg, P., and R. Li. 2003. Cell Cycle. 2:202-210; Ganem, N.J., Z. Storchova, and D. Pellman. 2007. Curr. Opin. Genet. Dev. 17:157-162). However, there is little direct evidence linking cancer-causing mutations with such events, and it remains controversial whether genomic instability is a cause or an effect of cancer. In this study, we show that adenomatous polyposis coli (APC) mutations found in human colorectal cancers dominantly inhibit cytokinesis by preventing mitotic spindle anchoring at the anaphase cortex and, thus, blocking initiation of the cytokinetic furrow. We find that dividing crypt cells in the small intestines of APC(Min/+) mice exhibit similar mitotic defects, including misoriented spindles and misaligned chromosomes. These defects are observed in normal crypt cells with wild-type levels of beta-catenin and, importantly, are associated with tetraploid genotypes. We provide direct evidence that the dominant activity of APC mutants induces aneuploidy in vivo. Our data support a model whereby tetraploid cells represent a first step in the onset of genomic instability and colorectal cancer.
Figures





References
-
- Cardoso, J., L. Molenaar, R.X. de Menezes, M. van Leerdam, C. Rosenberg, G. Moslein, J. Sampson, H. Morreau, J.M. Boer, and R. Fodde. 2006. Chromosomal instability in MYH- and APC-mutant adenomatous polyps. Cancer Res. 66:2514–2519. - PubMed
-
- D'Avino, P.P., M.S. Savoian, and D.M. Glover. 2005. Cleavage furrow formation and ingression during animal cytokinesis: a microtubule legacy. J. Cell Sci. 118:1549–1558. - PubMed
-
- Duesberg, P., and R. Li. 2003. Multistep carcinogenesis: a chain reaction of aneuploidizations. Cell Cycle. 2:202–210. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
