

Actin-myosin network reorganization breaks symmetry at the cell rear to spontaneously initiate polarized cell motility
- PMID: 17893245
- PMCID: PMC2064654
- DOI: 10.1083/jcb.200706012
Actin-myosin network reorganization breaks symmetry at the cell rear to spontaneously initiate polarized cell motility
Abstract
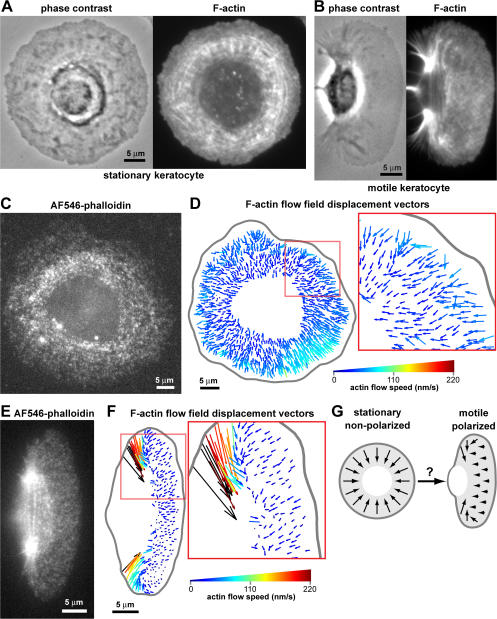
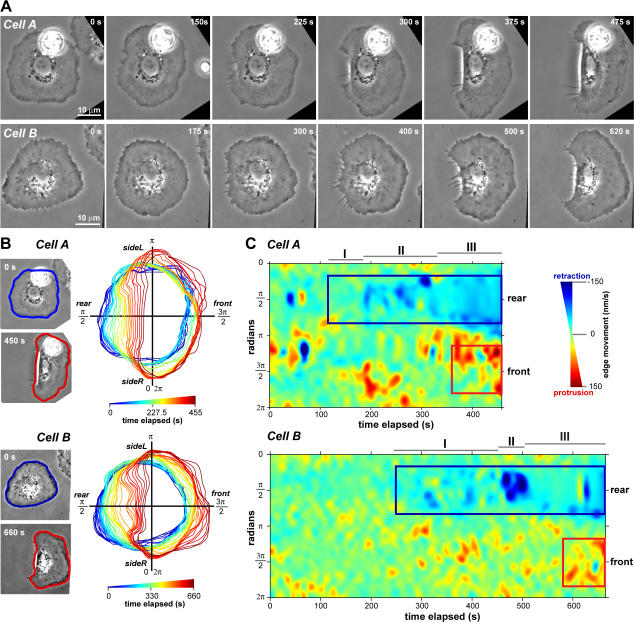
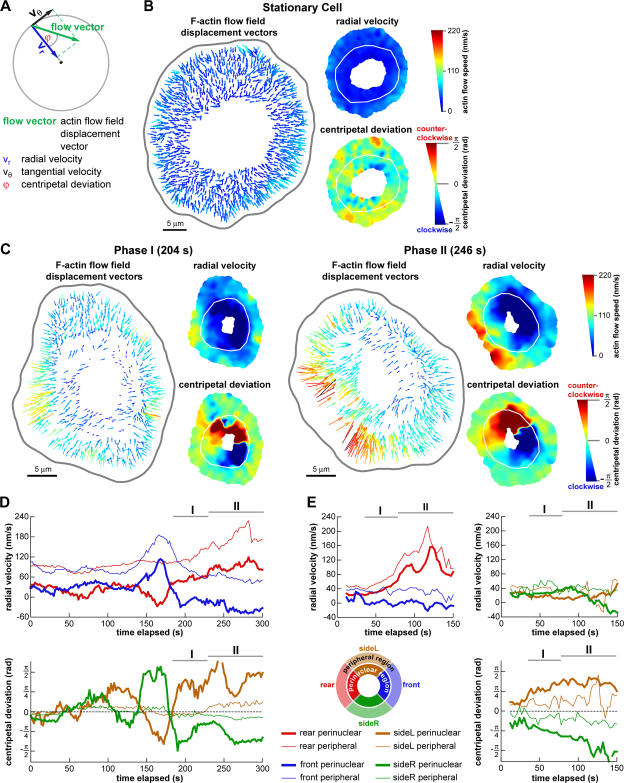
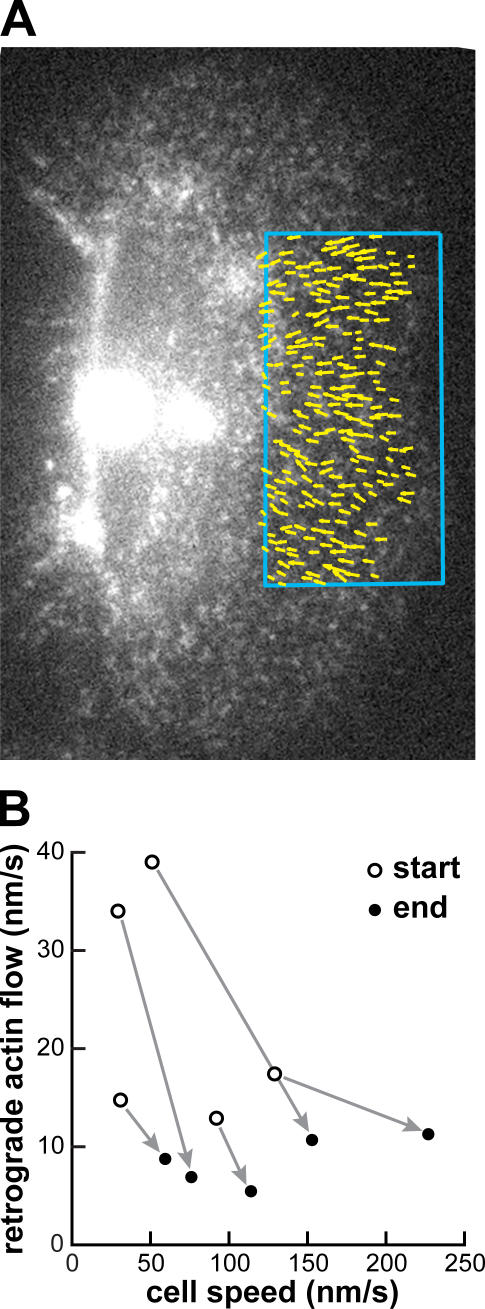
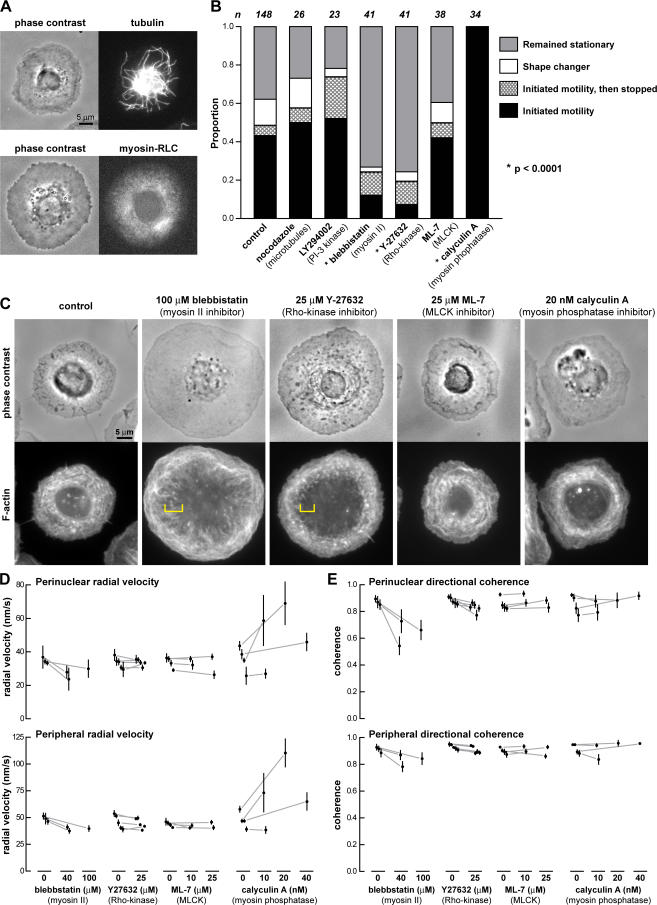
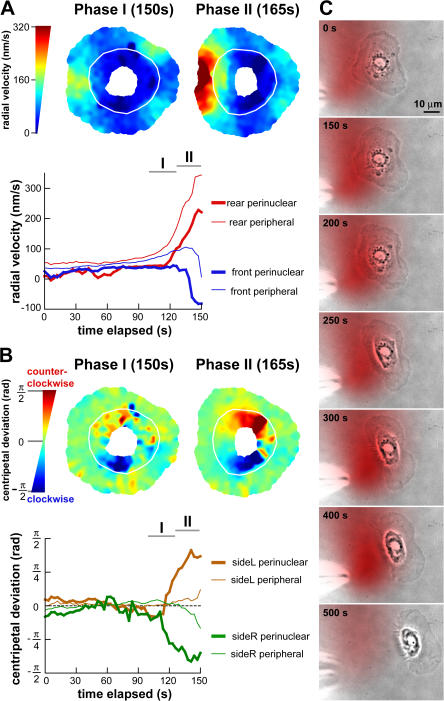
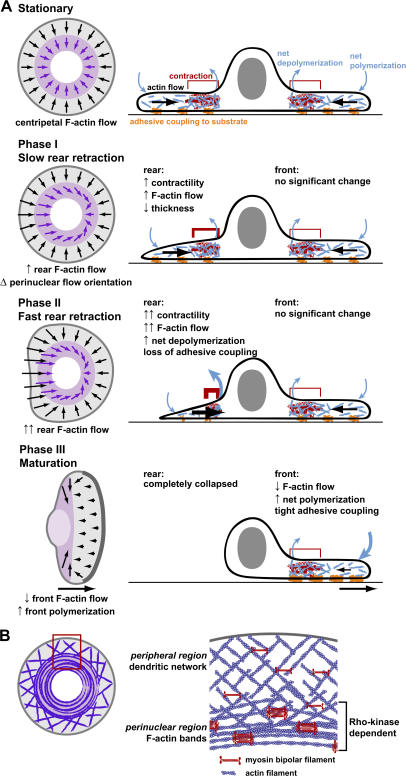
We have analyzed the spontaneous symmetry breaking and initiation of actin-based motility in keratocytes (fish epithelial cells). In stationary keratocytes, the actin network flow was inwards and radially symmetric. Immediately before motility initiation, the actin network flow increased at the prospective cell rear and reoriented in the perinuclear region, aligning with the prospective axis of movement. Changes in actin network flow at the cell front were detectable only after cell polarization. Inhibition of myosin II or Rho kinase disrupted actin network organization and flow in the perinuclear region and decreased the motility initiation frequency, whereas increasing myosin II activity with calyculin A increased the motility initiation frequency. Local stimulation of myosin activity in stationary cells by the local application of calyculin A induced directed motility initiation away from the site of stimulation. Together, these results indicate that large-scale actin-myosin network reorganization and contractility at the cell rear initiate spontaneous symmetry breaking and polarized motility of keratocytes.
Figures
References
-
- Dunn, G., and D. Zicha. 1995. Dynamics of fibroblast spreading. J. Cell Sci. 108:1239–1249. - PubMed
