

The type III effector EspF coordinates membrane trafficking by the spatiotemporal activation of two eukaryotic signaling pathways
- PMID: 17893247
- PMCID: PMC2064658
- DOI: 10.1083/jcb.200705021
The type III effector EspF coordinates membrane trafficking by the spatiotemporal activation of two eukaryotic signaling pathways
Abstract
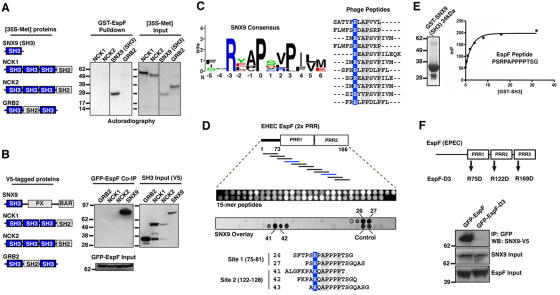
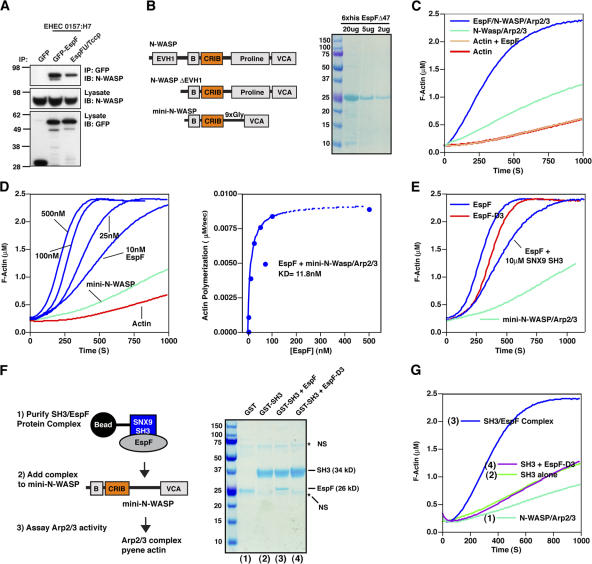
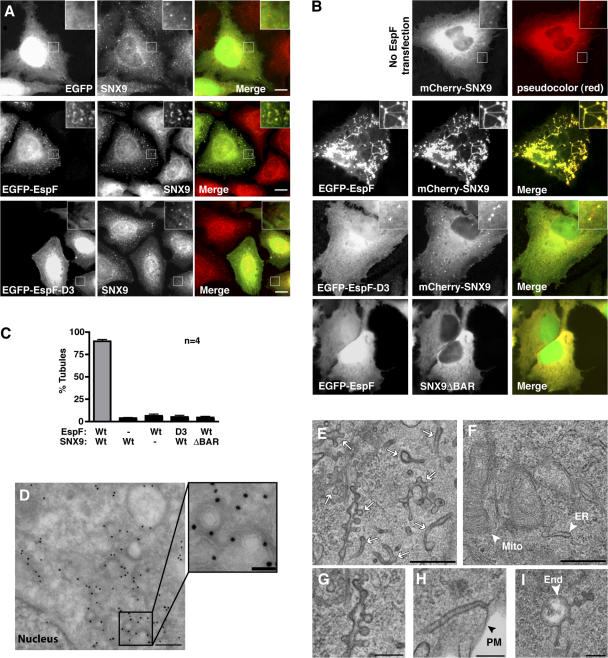
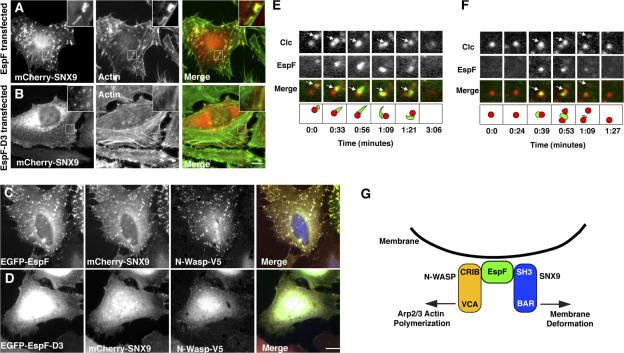
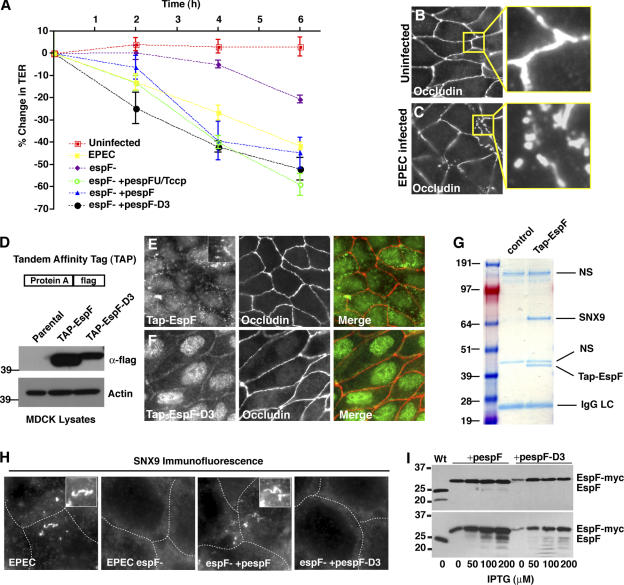
Bacterial toxins and effector proteins hijack eukaryotic enzymes that are spatially localized and display rapid signaling kinetics. However, the molecular mechanisms by which virulence factors engage highly dynamic substrates in the host cell environment are poorly understood. Here, we demonstrate that the enteropathogenic Escherichia coli (EPEC) type III effector protein EspF nucleates a multiprotein signaling complex composed of eukaryotic sorting nexin 9 (SNX9) and neuronal Wiskott-Aldrich syndrome protein (N-WASP). We demonstrate that a specific and high affinity association between EspF and SNX9 induces membrane remodeling in host cells. These membrane-remodeling events are directly coupled to N-WASP/Arp2/3-mediated actin nucleation. In addition to providing a biochemical mechanism of EspF function, we find that EspF dynamically localizes to membrane-trafficking organelles in a spatiotemporal pattern that correlates with SNX9 and N-WASP activity in living cells. Thus, our findings suggest that the EspF-dependent assembly of SNX9 and N-WASP represents a novel form of signaling mimicry used to promote EPEC pathogenesis and gastrointestinal disease.
Figures


References
-
- Alto, N.M., F. Shao, C.S. Lazar, R.L. Brost, G. Chua, S. Mattoo, S.A. McMahon, P. Ghosh, T.R. Hughes, C. Boone, and J.E. Dixon. 2006. Identification of a bacterial type III effector family with G protein mimicry functions. Cell. 124:133–145. - PubMed
-
- Badour, K., M.K. McGavin, J. Zhang, S. Freeman, C. Vieira, D. Filipp, M. Julius, G.B. Mills, and K.A. Siminovitch. 2007. Interaction of the Wiskott-Aldrich syndrome protein with sorting nexin 9 is required for CD28 endocytosis and cosignaling in T cells. Proc. Natl. Acad. Sci. USA. 104:1593–1598. - PMC - PubMed
-
- Benesch, S., S. Polo, F.P. Lai, K.I. Anderson, T.E. Stradal, J. Wehland, and K. Rottner. 2005. N-WASP deficiency impairs EGF internalization and actin assembly at clathrin-coated pits. J. Cell Sci. 118:3103–3115. - PubMed
-
- Campellone, K.G., D. Robbins, and J.M. Leong. 2004. EspFU is a translocated EHEC effector that interacts with Tir and N-WASP and promotes Nck-independent actin assembly. Dev. Cell. 7:217–228. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
