

Time-resolved NMR methods resolving ligand-induced RNA folding at atomic resolution
- PMID: 17895388
- PMCID: PMC2000436
- DOI: 10.1073/pnas.0703182104
Time-resolved NMR methods resolving ligand-induced RNA folding at atomic resolution
Abstract
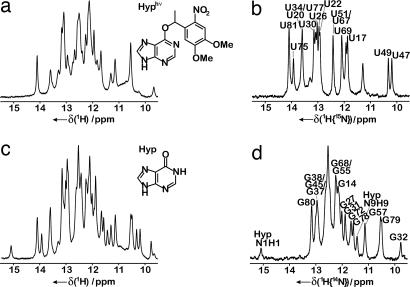
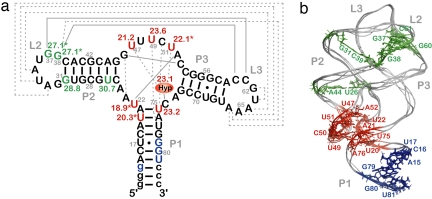
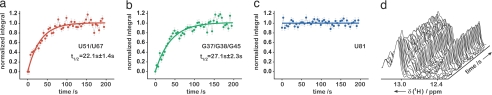
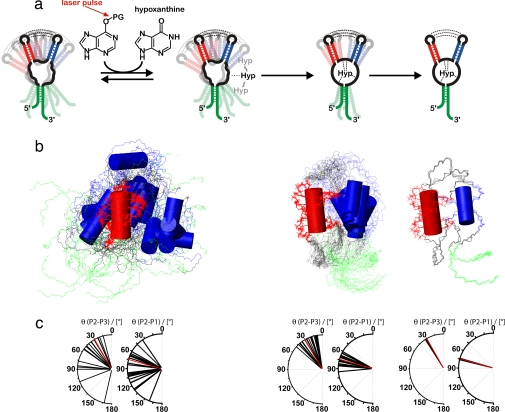
Structural transitions of RNA between alternate conformations with similar stabilities are associated with important aspects of cellular function. Few techniques presently exist that are capable of monitoring such transitions and thereby provide insight into RNA dynamics and function at atomic resolution. Riboswitches are found in the 5'-UTR of mRNA and control gene expression through structural transitions after ligand recognition. A time-resolved NMR strategy was established in conjunction with laser-triggered release of the ligand from a photocaged derivative in situ to monitor the hypoxanthine-induced folding of the guanine-sensing riboswitch aptamer domain of the Bacillus subtilis xpt-pbuX operon at atomic resolution. Combining selective isotope labeling of the RNA with NMR filter techniques resulted in significant spectral resolution and allowed kinetic analysis of the buildup rates for individual nucleotides in real time. Three distinct kinetic steps associated with the ligand-induced folding were delineated. After initial complex encounter the ligand-binding pocket is formed and results in subsequent stabilization of a remote long-range loop-loop interaction. Incorporation of NMR data into experimentally restrained molecular dynamics simulations provided insight into the RNA structural ensembles involved during the conformational transition.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Radford SE, Dobson CM, Evans PA. Nature. 1992;358:302–307. - PubMed
-
- Vendruscolo M, Paci E, Dobson CM, Karplus M. Nature. 2001;409:641–645. - PubMed
-
- Wyatt JR, Puglisi JD, Tinoco I., Jr BioEssays. 1989;11:100–106. - PubMed
-
- Schroeder R, Barta A, Semrad K. Nat Rev Mol Cell Biol. 2004;5:908–919. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
