

Do individual females differ intrinsically in their propensity to engage in extra-pair copulations?
- PMID: 17895992
- PMCID: PMC1978515
- DOI: 10.1371/journal.pone.0000952
Do individual females differ intrinsically in their propensity to engage in extra-pair copulations?
Abstract
Background: While many studies have investigated the occurrence of extra-pair paternity in wild populations of birds, we still know surprisingly little about whether individual females differ intrinsically in their principal readiness to copulate, and to what extent this readiness is affected by male attractiveness.
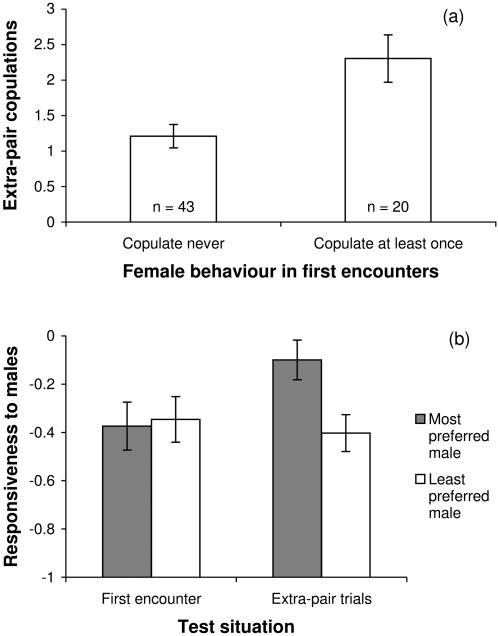
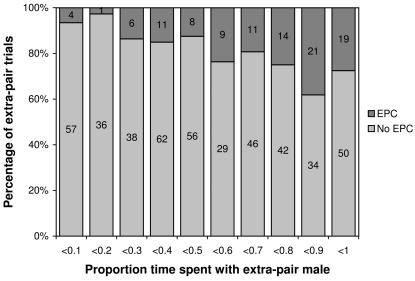
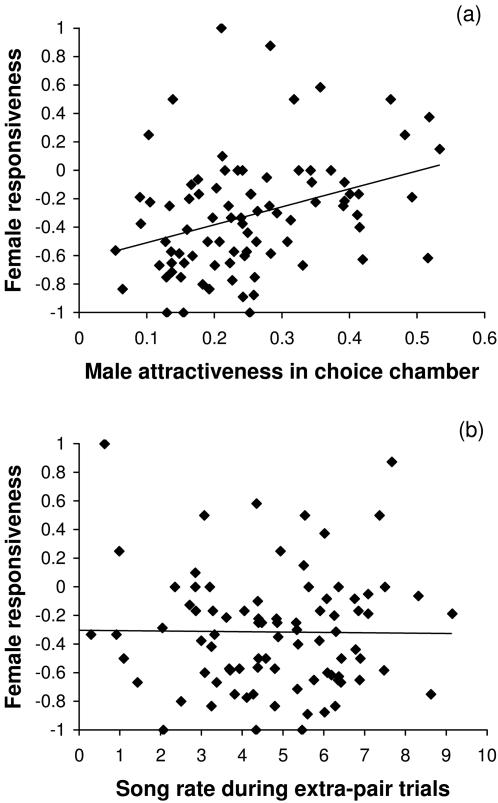
Methodology/findings: To address this question I used captive zebra finches (Taeniopygia guttata) as a model system. I first measured female readiness to copulate when courted by a male for the first time in life. Second, I conducted choice-chamber experiments to assess the mating preferences of individual females prior to pair formation. I then paired females socially with a non-desired mate and once they had formed a stable pair bond, I observed the inclination of these females to engage in extra-pair copulations with various males. Females showing a high readiness to copulate when courted by a male for the first time in life were much more likely to engage in extra-pair copulations later in life than others. Male attractiveness, as measured in choice tests, was a useful predictor of whether females engaged in extra-pair copulations with these males, but, surprisingly, the attractiveness of a female's social partner had no effect on her fidelity. However, it remained unclear what made some males more attractive than others. Contrary to a widespread but rarely tested hypothesis, females did not preferentially copulate with males having a redder beak or singing at a higher rate. Rather it seemed that song rate was a confounding factor in choice-chamber experiments: song attracted the female's attention but did not increase the male's attractiveness as a copulation partner.
Conclusions/significance: Intrinsic variation in female readiness to copulate as well as variation in the attractiveness of the extra-pair male but not the social partner decided the outcome of extra-pair encounters.
Conflict of interest statement
Figures
References
-
- Birkhead TR, Møller AP. London: Academic Press; 1992. Sperm competition in birds.
-
- Petrie M, Kempenaers B. Extra-pair paternity in birds: explaining variation between species and populations. Trends Ecol Evol. 1998;13:52–58. - PubMed
-
- Griffith SC, Owens IPF, Thuman KA. Extra pair paternity in birds: a review of interspecific variation and adaptive function. Mol Ecol. 2002;11:2195–2212. - PubMed
-
- Westneat DF, Stewart IRK. Extra-pair paternity in birds: causes, correlates, and conflict. Ann Rev Ecol Evol Syst. 2003;34:365–396.
-
- Johnson K, Burley N. Mating tactics and mating systems of birds. In: Parker (eds. P. G. , Burley N. T. , editors. Avian Reproductive Tactics: Female and Male Perspectives. Ornithol Monogr. Vol. 49. 1998. pp. 1–20.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
