

MVB-12, a fourth subunit of metazoan ESCRT-I, functions in receptor downregulation
- PMID: 17895996
- PMCID: PMC1978536
- DOI: 10.1371/journal.pone.0000956
MVB-12, a fourth subunit of metazoan ESCRT-I, functions in receptor downregulation
Abstract
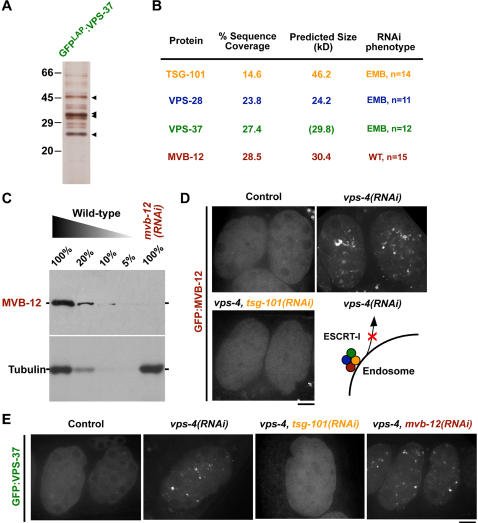
After ligand binding and endocytosis, cell surface receptors can continue to signal from endosomal compartments until sequestered from the cytoplasm. An important mechanism for receptor downregulation in vivo is via the inward budding of receptors into intralumenal vesicles to form specialized endosomes called multivesicular bodies (MVBs) that subsequently fuse with lysosomes, degrading their cargo. This process requires four heterooligomeric protein complexes collectively termed the ESCRT machinery. In yeast, ESCRT-I is a heterotetrameric complex comprised of three conserved subunits and a fourth subunit for which identifiable metazoan homologs were lacking. Using C. elegans, we identify MVB-12, a fourth metazoan ESCRT-I subunit. Depletion of MVB-12 slows the kinetics of receptor downregulation in vivo, but to a lesser extent than inhibition of other ESCRT-I subunits. Consistent with these findings, targeting of MVB-12 to membranes requires the other ESCRT-I subunits, but MVB-12 is not required to target the remaining ESCRT-I components. Both endogenous and recombinant ESCRT-I are stable complexes with a 1:1:1:1 subunit stoichiometry. MVB-12 has two human homologs that co-localize and co-immunoprecipitate with the ESCRT-I component TSG101. Thus, MVB-12 is a conserved core component of metazoan ESCRT-I that regulates its activity during MVB biogenesis.
Conflict of interest statement
Figures




References
-
- Schlessinger J. Cell signaling by receptor tyrosine kinases. Cell. 2000;103:211–225. - PubMed
-
- Vivekanand P, Rebay I. Intersection of signal transduction pathways and development. Annu Rev Genet. 2006;40:139–157. - PubMed
-
- Porter AC, Vaillancourt RR. Tyrosine kinase receptor-activated signal transduction pathways which lead to oncogenesis. Oncogene. 1998;17:1343–1352. - PubMed
-
- Katzmann DJ, Odorizzi G, Emr SD. Receptor downregulation and multivesicular-body sorting. Nat Rev Mol Cell Biol. 2002;3:893–905. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
