

Dyneins across eukaryotes: a comparative genomic analysis
- PMID: 17897317
- PMCID: PMC2239267
- DOI: 10.1111/j.1600-0854.2007.00646.x
Dyneins across eukaryotes: a comparative genomic analysis
Abstract
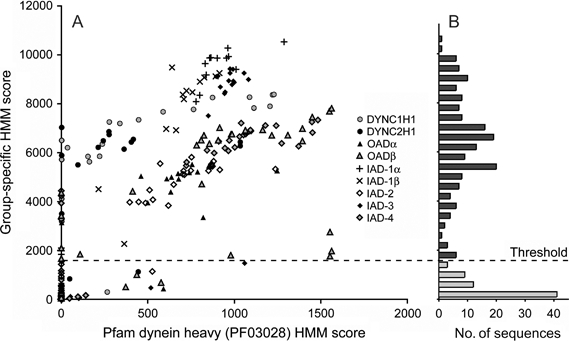
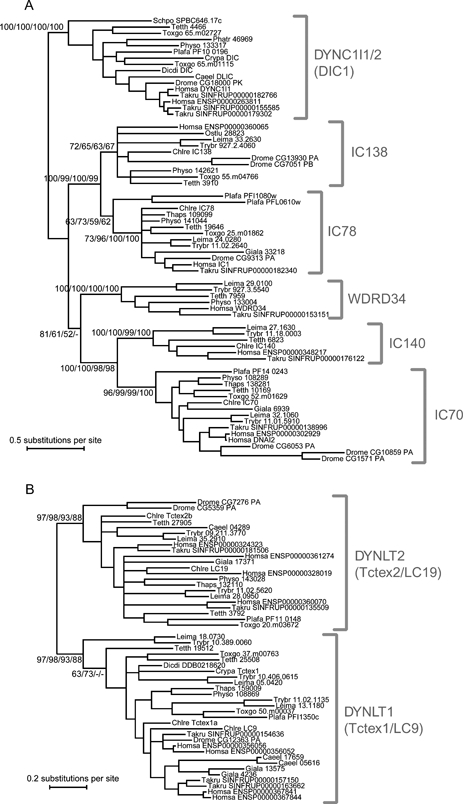
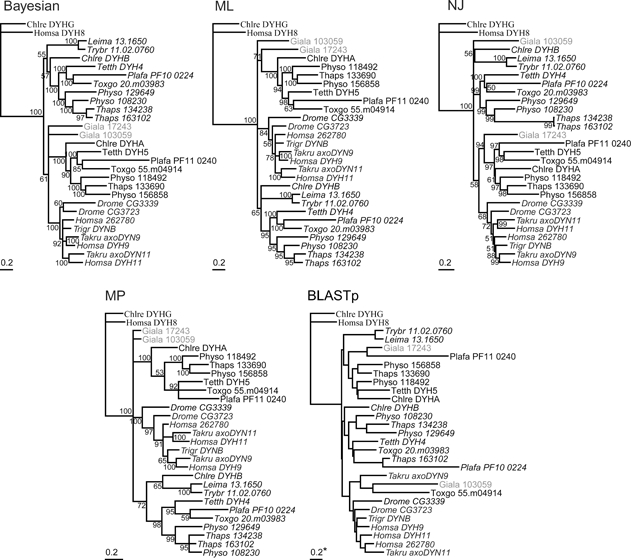
Dyneins are large minus-end-directed microtubule motors. Each dynein contains at least one dynein heavy chain (DHC) and a variable number of intermediate chains (IC), light intermediate chains (LIC) and light chains (LC). Here, we used genome sequence data from 24 diverse eukaryotes to assess the distribution of DHCs, ICs, LICs and LCs across Eukaryota. Phylogenetic inference identified nine DHC families (two cytoplasmic and seven axonemal) and six IC families (one cytoplasmic). We confirm that dyneins have been lost from higher plants and show that this is most likely because of a single loss of cytoplasmic dynein 1 from the ancestor of Rhodophyta and Viridiplantae, followed by lineage-specific losses of other families. Independent losses in Entamoeba mean that at least three extant eukaryotic lineages are entirely devoid of dyneins. Cytoplasmic dynein 2 is associated with intraflagellar transport (IFT), but in two chromalveolate organisms, we find an IFT footprint without the retrograde motor. The distribution of one family of outer-arm dyneins accounts for 2-headed or 3-headed outer-arm ultrastructures observed in different organisms. One diatom species builds motile axonemes without any inner-arm dyneins (IAD), and the unexpected conservation of IAD I1 in non-flagellate algae and LC8 (DYNLL1/2) in all lineages reveals a surprising fluidity to dynein function.
Figures


References
-
- Hirokawa N, Noda Y, Okada Y. Kinesin and dynein superfamily proteins in organelle transport and cell division. Curr Opin Cell Biol. 1998;10:60–73. - PubMed
-
- Gibbons IR. The role of dynein in microtubule-based motility. Cell Struct Funct. 1996;21:331–342. - PubMed
-
- Holzbaur ELF, Vallee RB. Dyneins: molecular structure and cellular function. Annu Rev Cell Biol. 1994;10:339–372. - PubMed
-
- Kon T, Nishiura M, Ohkura R, Toyoshima YY, Sutoh K. Distinct functions of nucleotide-binding/hydrolysis sites in the four AAA modules of cytoplasmic dynein. Biochemistry. 2004;43:11266–11274. - PubMed
-
- Gibbons IR, LeeEiford A, Mocz G, Phillipson CA, Tang WJY, Gibbons BH. Photosensitized cleavage of dynein heavy chains: cleavage at the V1 site by irradiation at 365nm in the presence of ATP and vanadate. J Biol Chem. 1987;262:2780–2786. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
