

Origin and distribution of epipolythiodioxopiperazine (ETP) gene clusters in filamentous ascomycetes
- PMID: 17897469
- PMCID: PMC2045112
- DOI: 10.1186/1471-2148-7-174
Origin and distribution of epipolythiodioxopiperazine (ETP) gene clusters in filamentous ascomycetes
Abstract
Background: Genes responsible for biosynthesis of fungal secondary metabolites are usually tightly clustered in the genome and co-regulated with metabolite production. Epipolythiodioxopiperazines (ETPs) are a class of secondary metabolite toxins produced by disparate ascomycete fungi and implicated in several animal and plant diseases. Gene clusters responsible for their production have previously been defined in only two fungi. Fungal genome sequence data have been surveyed for the presence of putative ETP clusters and cluster data have been generated from several fungal taxa where genome sequences are not available. Phylogenetic analysis of cluster genes has been used to investigate the assembly and heredity of these gene clusters.
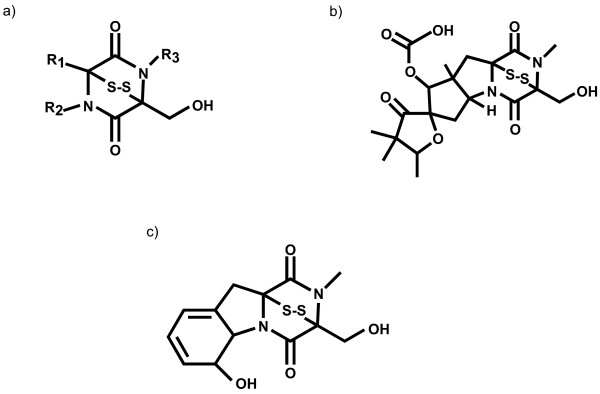
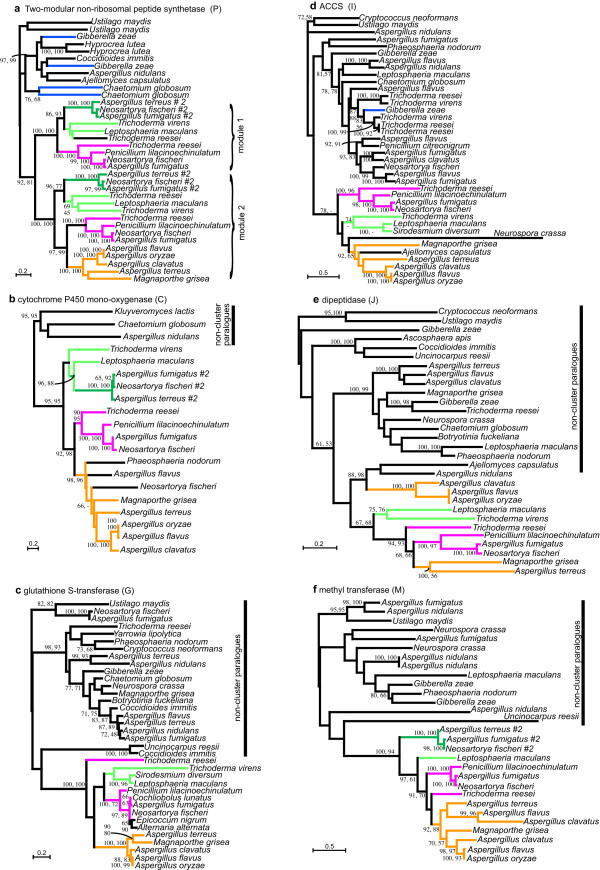
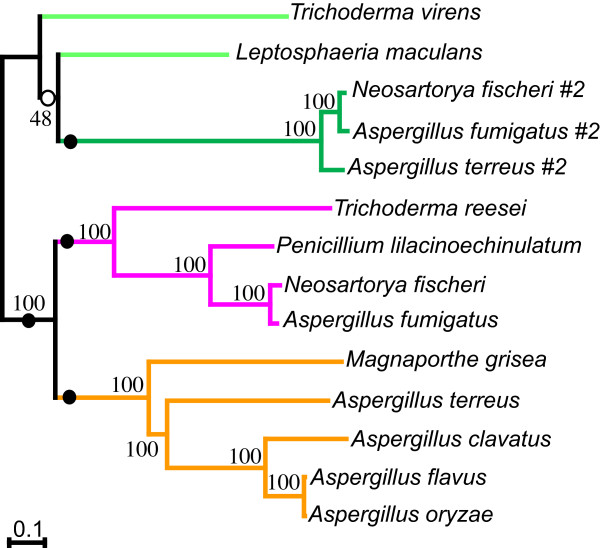
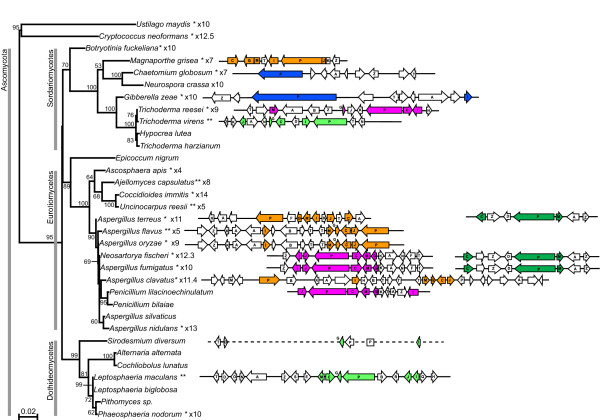
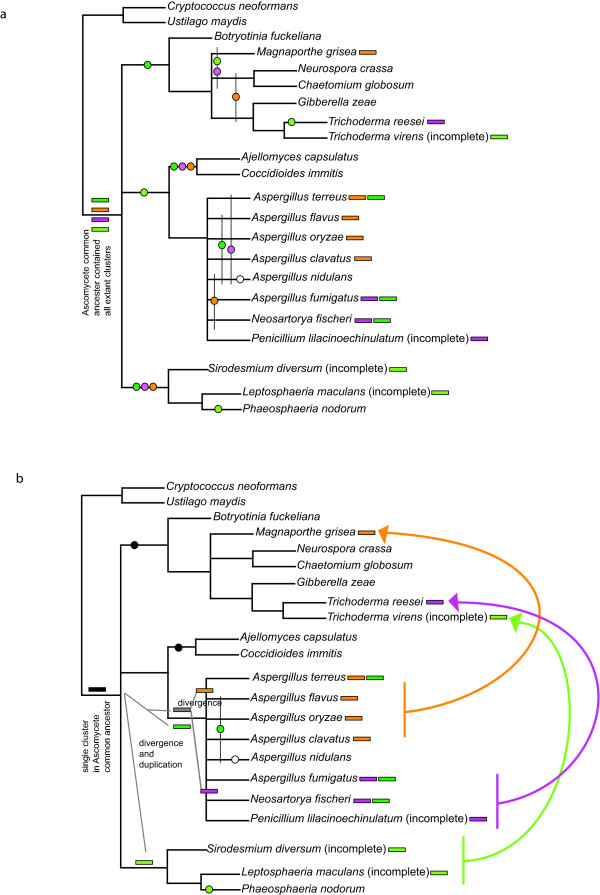
Results: Putative ETP gene clusters are present in 14 ascomycete taxa, but absent in numerous other ascomycetes examined. These clusters are discontinuously distributed in ascomycete lineages. Gene content is not absolutely fixed, however, common genes are identified and phylogenies of six of these are separately inferred. In each phylogeny almost all cluster genes form monophyletic clades with non-cluster fungal paralogues being the nearest outgroups. This relatedness of cluster genes suggests that a progenitor ETP gene cluster assembled within an ancestral taxon. Within each of the cluster clades, the cluster genes group together in consistent subclades, however, these relationships do not always reflect the phylogeny of ascomycetes. Micro-synteny of several of the genes within the clusters provides further support for these subclades.
Conclusion: ETP gene clusters appear to have a single origin and have been inherited relatively intact rather than assembling independently in the different ascomycete lineages. This progenitor cluster has given rise to a small number of distinct phylogenetic classes of clusters that are represented in a discontinuous pattern throughout ascomycetes. The disjunct heredity of these clusters is discussed with consideration to multiple instances of independent cluster loss and lateral transfer of gene clusters between lineages.
Figures

References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
