

Chronic ethanol and withdrawal differentially modulate pre- and postsynaptic function at glutamatergic synapses in rat basolateral amygdala
- PMID: 17898152
- PMCID: PMC2269699
- DOI: 10.1152/jn.00189.2007
Chronic ethanol and withdrawal differentially modulate pre- and postsynaptic function at glutamatergic synapses in rat basolateral amygdala
Abstract
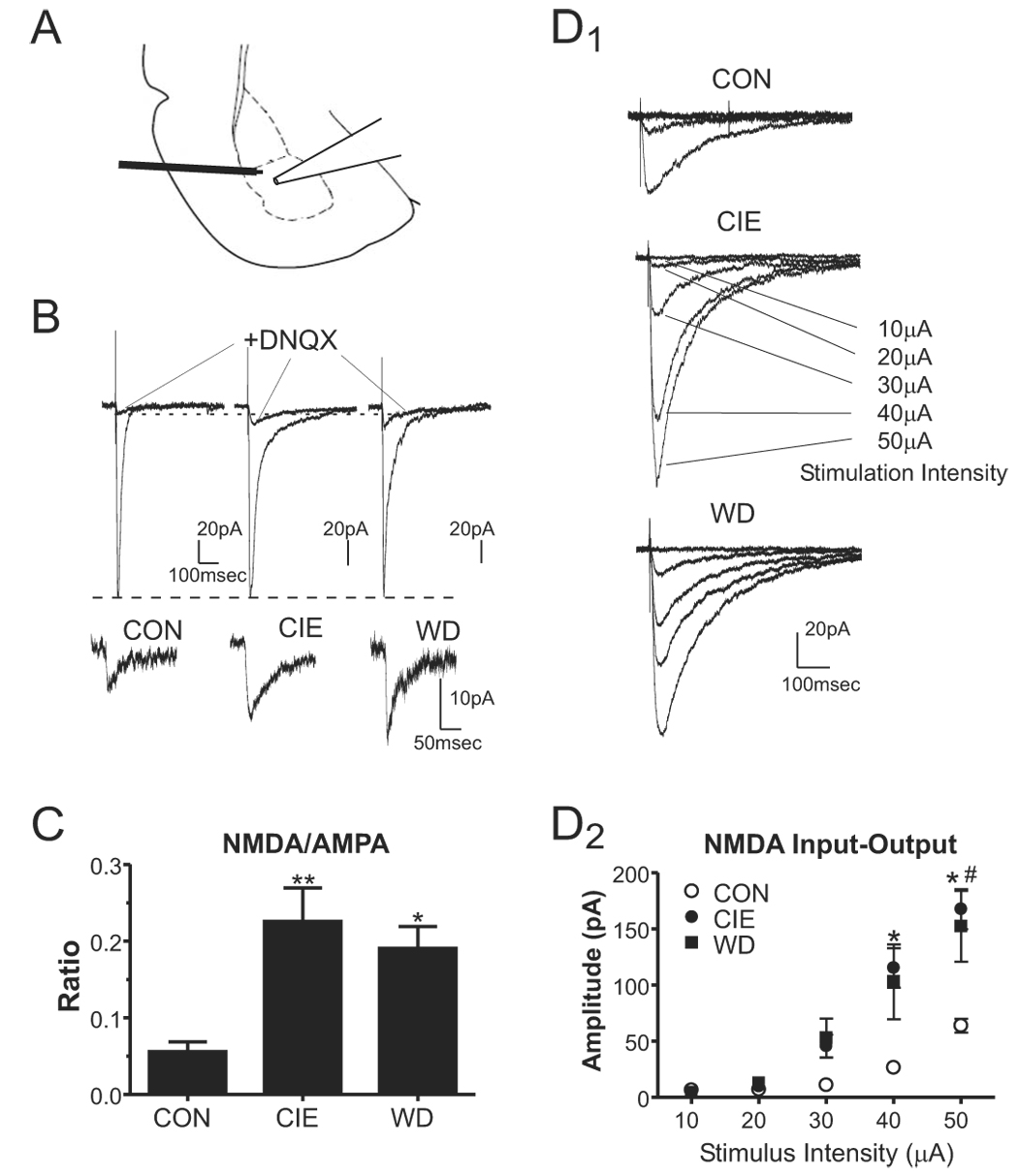
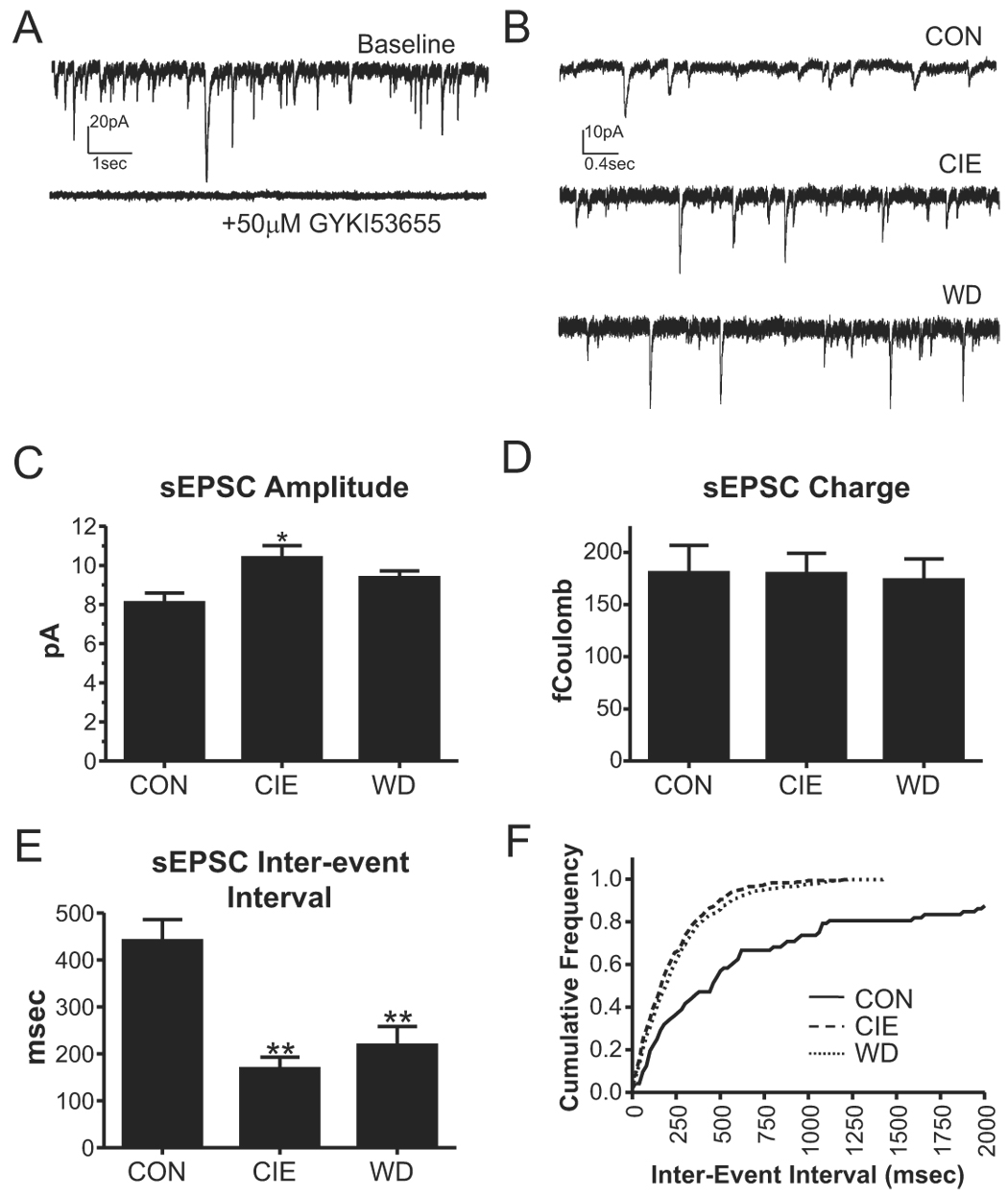
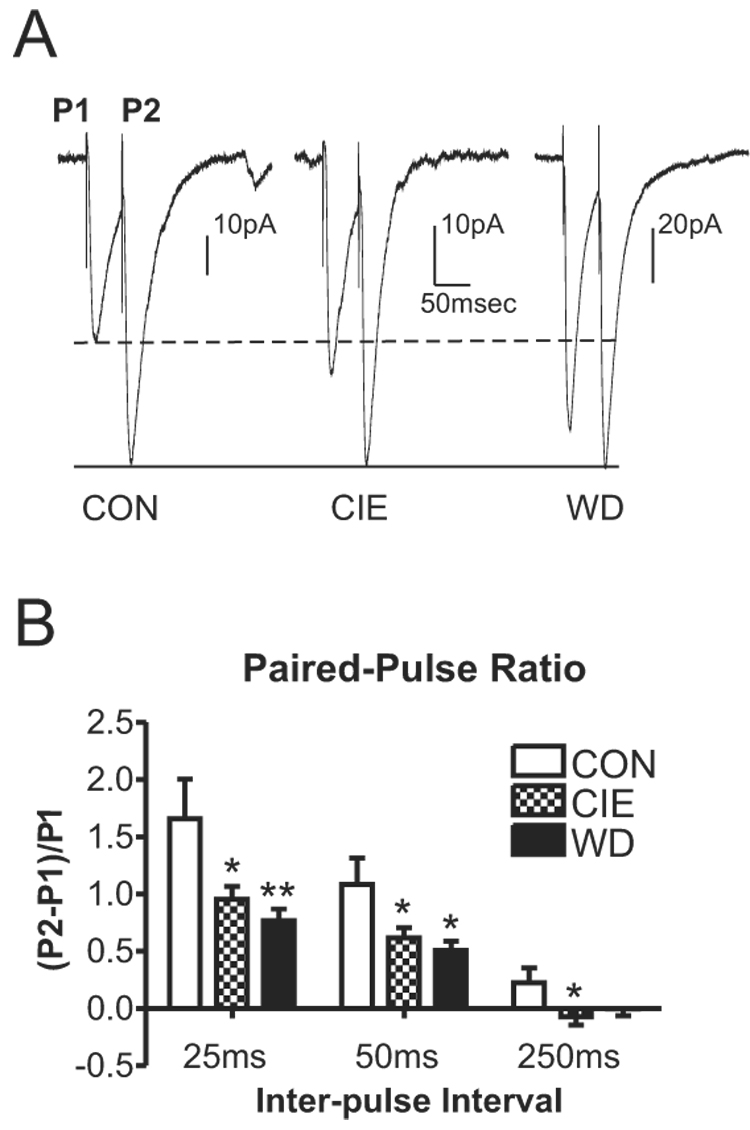
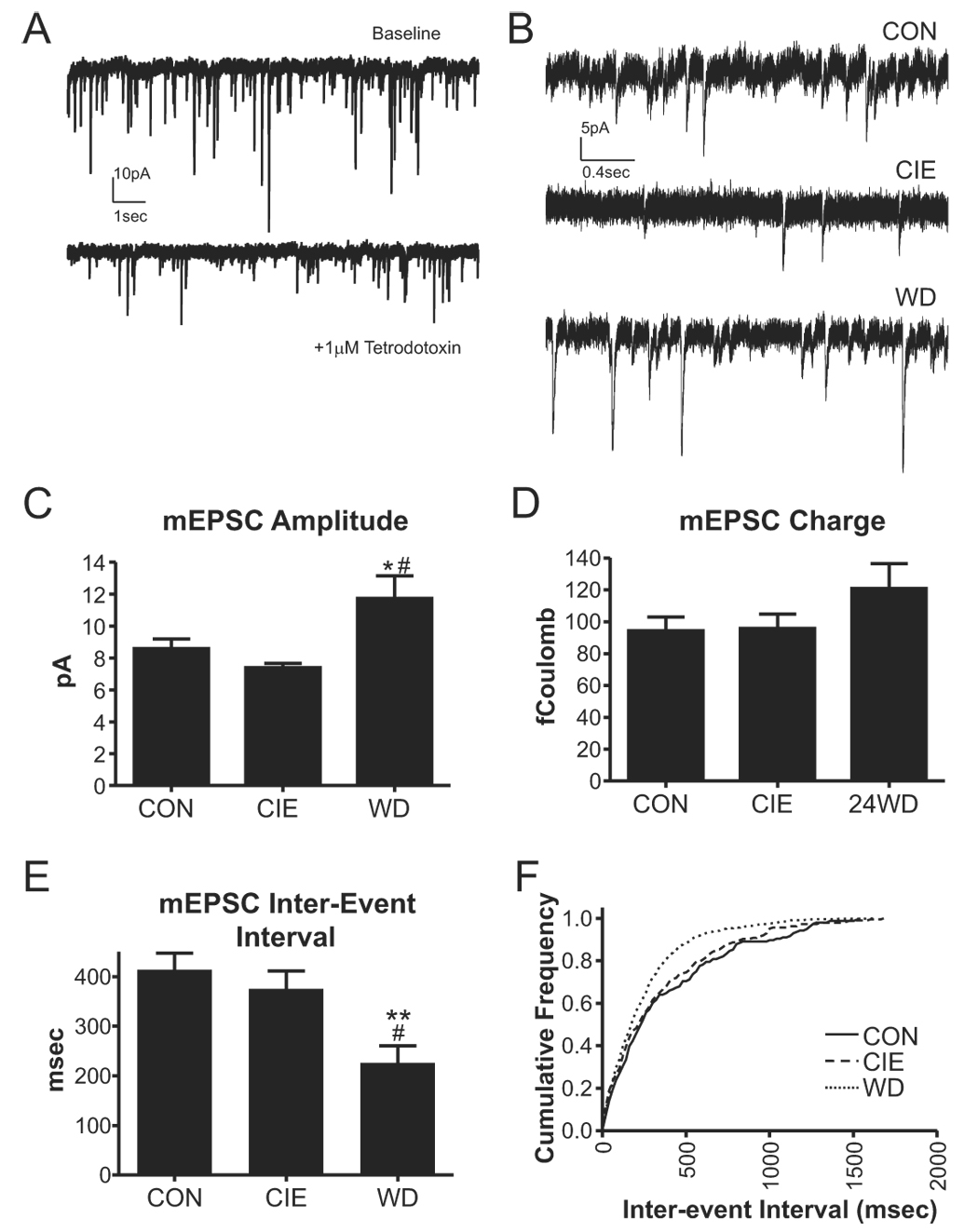
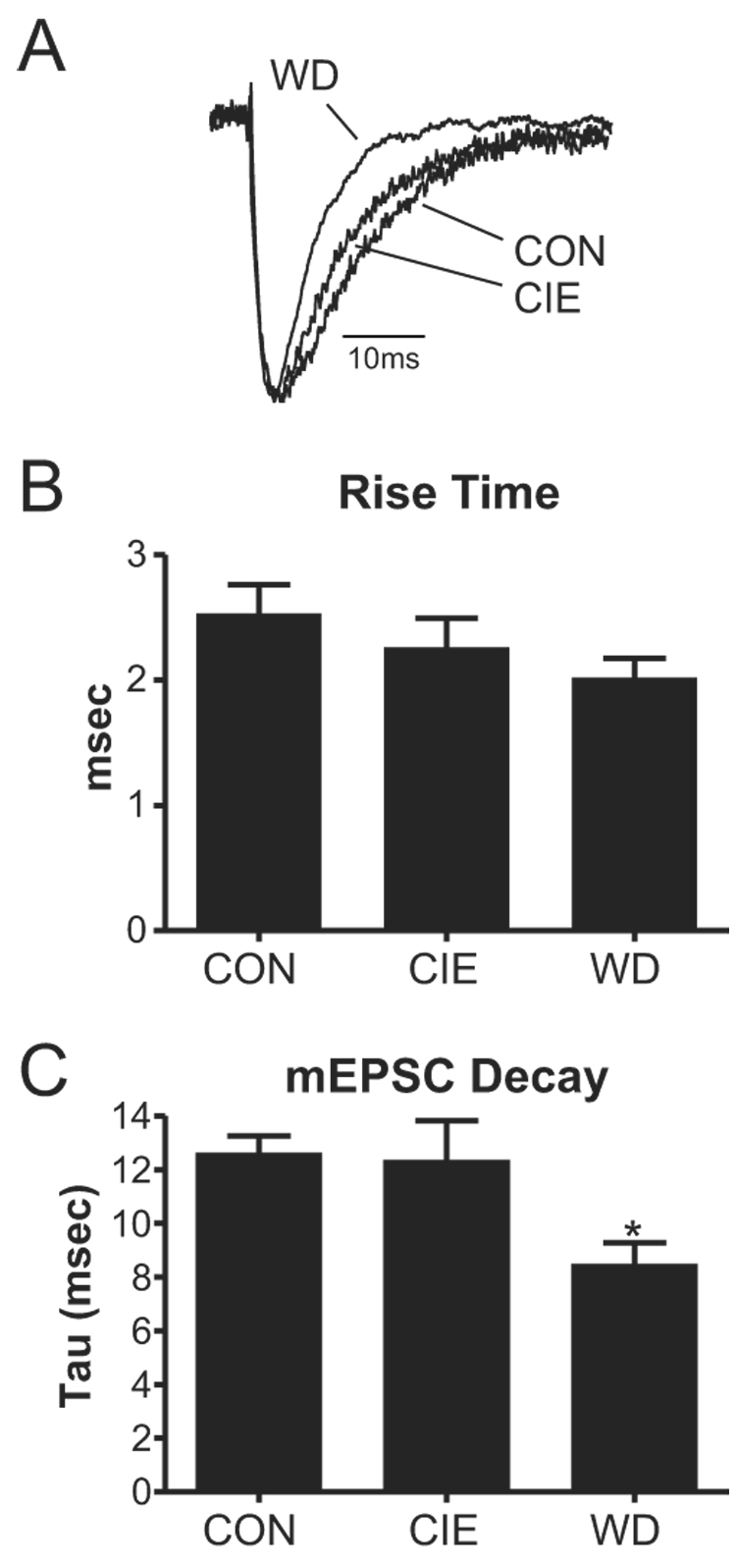
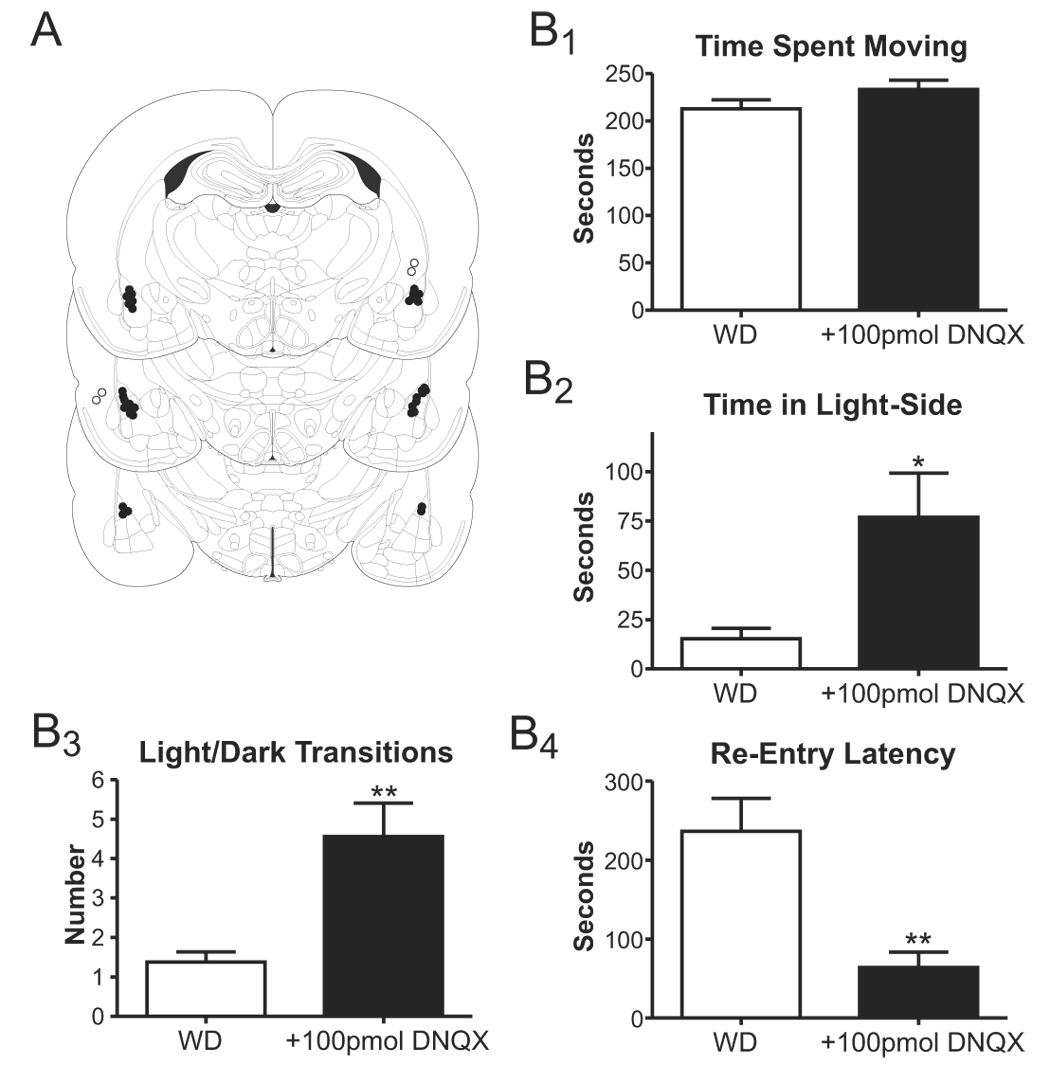
Withdrawal anxiety is a significant factor contributing to continued alcohol abuse in alcoholics. This anxiety is long-lasting, can manifest well after the overt physical symptoms of withdrawal, and is frequently associated with relapse in recovering alcoholics. The neurobiological mechanisms governing these withdrawal-associated increases in anxiety are currently unknown. The basolateral amygdala (BLA) is a major emotional center in the brain and regulates the expression of both learned fear and anxiety. Neurotransmitter system alterations within this brain region may therefore contribute to withdrawal-associated anxiety. Because evidence suggests that glutamate-gated neurotransmitter receptors are sensitive to acute ethanol exposure, we examined the effect of chronic intermittent ethanol (CIE) and withdrawal (WD) on glutamatergic synaptic transmission in the BLA. We found that slices prepared from CIE and WD animals had significantly increased contributions by synaptic NMDA receptors. In addition, CIE increased the amplitude of AMPA-receptor-mediated spontaneous excitatory postsynaptic currents (sEPSCs), whereas only WD altered the amplitude and kinetics of tetrodotoxin-resistant spontaneous events (mEPSCs). Similarly, the frequency of sEPSCs was increased in both CIE and WD neurons, although only WD increased the frequency of mEPSCs. These data suggest that CIE and WD differentially alter both pre- and postsynaptic properties of BLA glutamatergic synapses. Finally, we show that microinjection of the AMPA-receptor antagonist, DNQX, can attenuate withdrawal-related anxiety-like behavior. Together, our results suggest that increased glutamatergic function may contribute to anxiety expressed during withdrawal from chronic ethanol.
Figures
References
-
- Alderson HL, Robbins TW, Everitt BJ. The effects of excitotoxic lesions of the basolateral amygdala on the acquisition of heroin-seeking behaviour in rats. Psychopharmacology (Berl) 2000;153:111–119. - PubMed
-
- Andreasen M, Hablitz JJ. Paired-pulse facilitation in the dentate gyrus: a patch-clamp study in rat hippocampus in vitro. J Neurophysiol. 1994;72:326–336. - PubMed
-
- Atassi B, Glavinovic MI. Effect of cyclothiazide on spontaneous miniature excitatory postsynaptic currents in rat hippocampal pyramidal cells. Pflugers Arch. 1999;437:471–478. - PubMed
-
- Bacci A, Coco S, Pravettoni E, Schenk U, Armano S, Frassoni C, Verderio C, De Camilli P, Matteoli M. Chronic blockade of glutamate receptors enhances presynaptic release and downregulates the interaction between synaptophysin-synaptobrevin-vesicle-associated membrane protein 2. J Neurosci. 2001;21:6588–6596. - PMC - PubMed
-
- Becker HC, Hale RL. Repeated episodes of ethanol withdrawal potentiate the severity of subsequent withdrawal seizures: an animal model of alcohol withdrawal "kindling". Alcohol Clin Exp Res. 1993;17:94–98. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources