

What a difference a decade makes: insights into translesion DNA synthesis
- PMID: 17898175
- PMCID: PMC2000391
- DOI: 10.1073/pnas.0704219104
What a difference a decade makes: insights into translesion DNA synthesis
Abstract
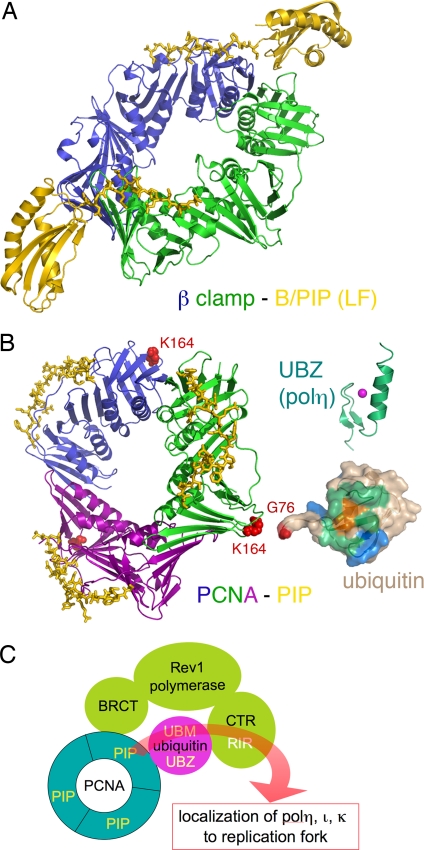
Living organisms are continually under attack from a vast array of DNA-damaging agents that imperils their genomic integrity. As a consequence, cells possess an army of enzymes to repair their damaged chromosomes. However, DNA lesions often persist and pose a considerable threat to survival, because they can block the cell's replicase and its ability to complete genome duplication. It has been clear for many years that cells must possess a mechanism whereby the DNA lesion could be tolerated and physically bypassed. Yet it was only within the past decade that specialized DNA polymerases for "translesion DNA synthesis" or "TLS" were identified and characterized. Many of the TLS enzymes belong to the recently described "Y-family" of DNA polymerases. By possessing a spacious preformed active site, these enzymes can physically accommodate a variety of DNA lesions and facilitate their bypass. Flexible DNA-binding domains and a variable binding pocket for the replicating base pair further allow these TLS polymerases to select specific lesions to bypass and favor distinct non-Watson-Crick base pairs. Consequently, TLS polymerases tend to exhibit much lower fidelity than the cell's replicase when copying normal DNA, which results in a dramatic increase in mutagenesis. Occasionally this can be beneficial, but it often speeds the onset of cancer in humans. Cells use both transcriptional and posttranslational regulation to keep these low-fidelity polymerases under strict control and limit their access to a replication fork. Our perspective focuses on the mechanistic insights into TLS by the Y-family polymerases, how they are regulated, and their effects on genomic (in)stability that have been described in the past decade.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
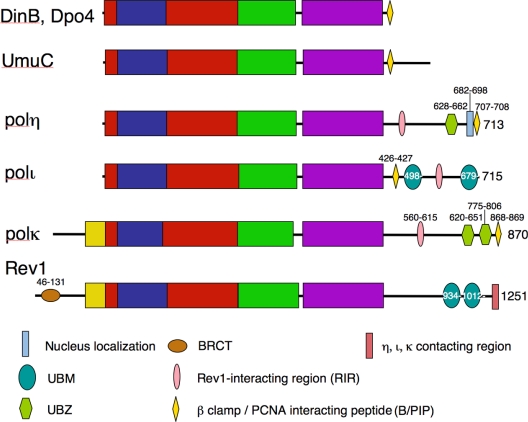
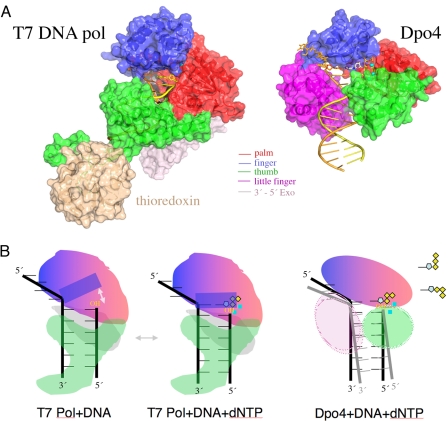
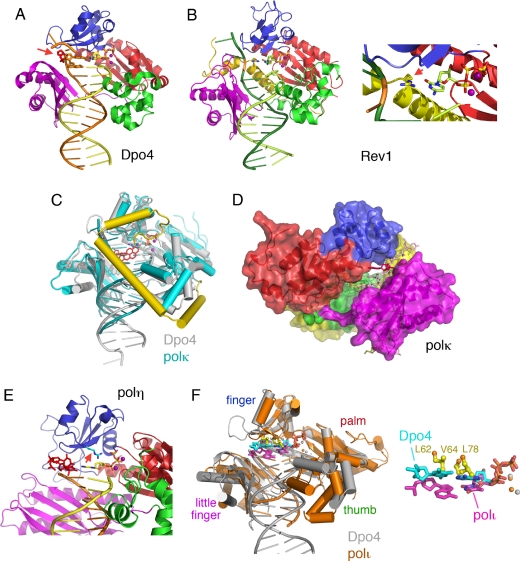
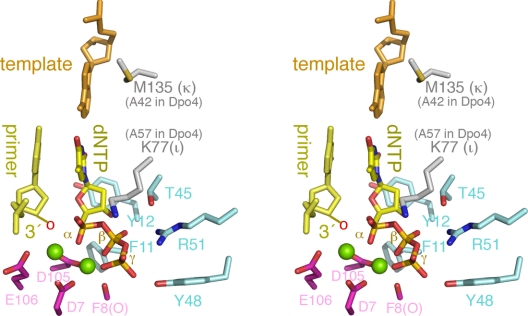
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
