

Auditory cortical receptive fields: stable entities with plastic abilities
- PMID: 17898209
- PMCID: PMC6673154
- DOI: 10.1523/JNEUROSCI.1462-07.2007
Auditory cortical receptive fields: stable entities with plastic abilities
Abstract
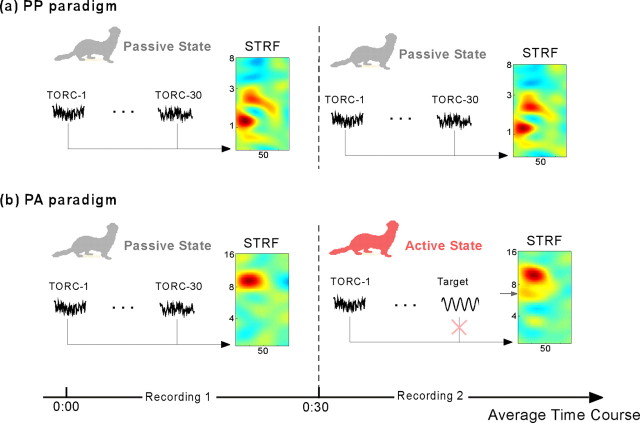
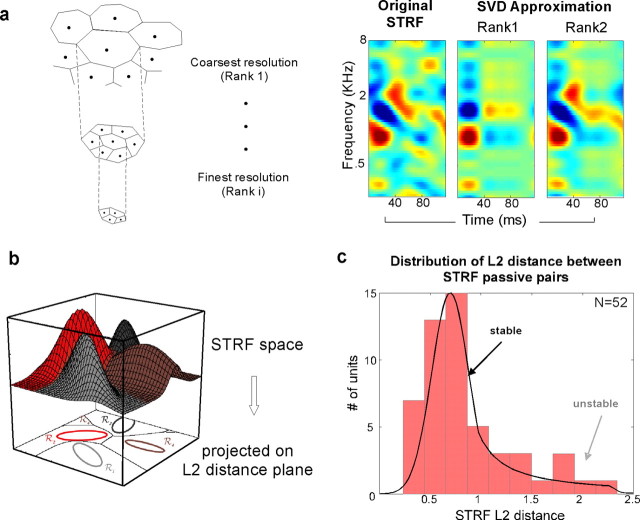
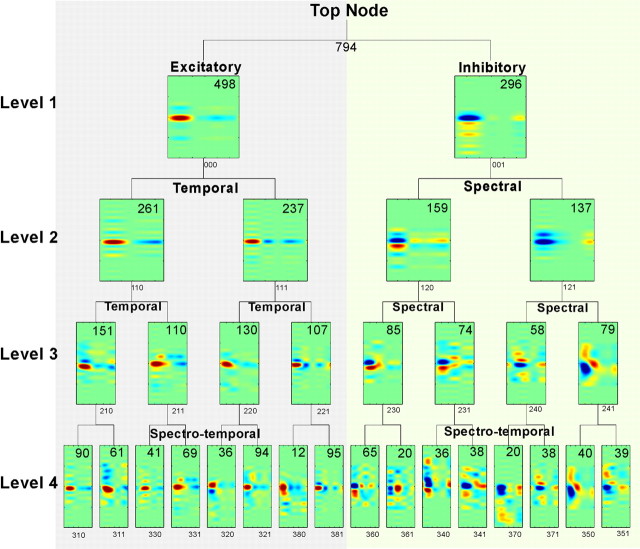
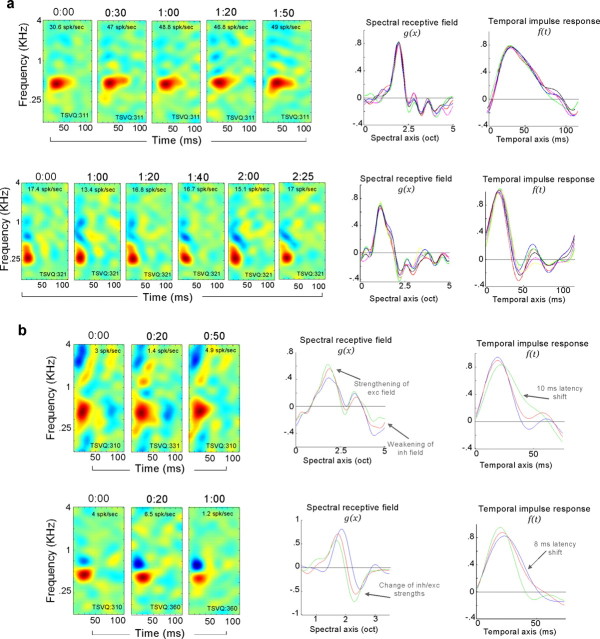
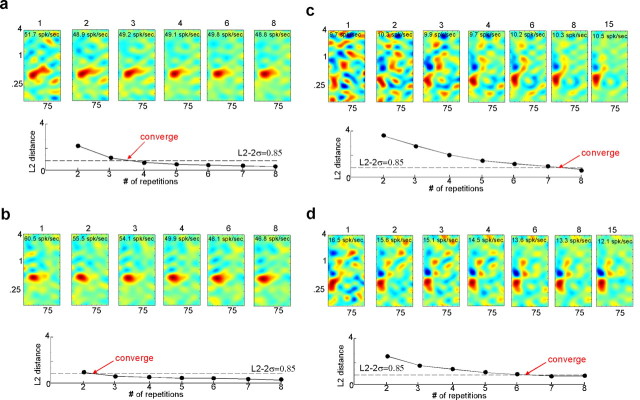
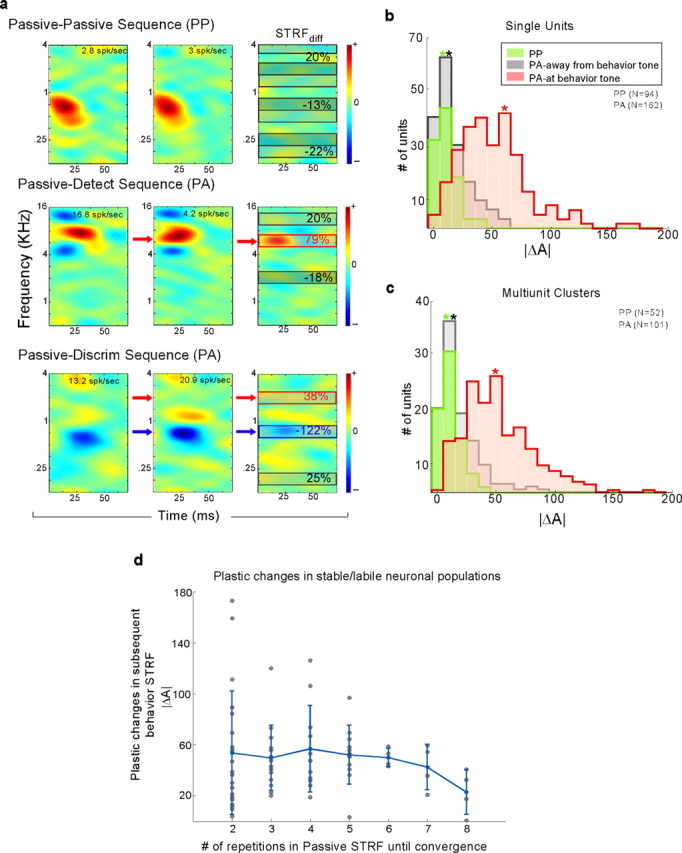
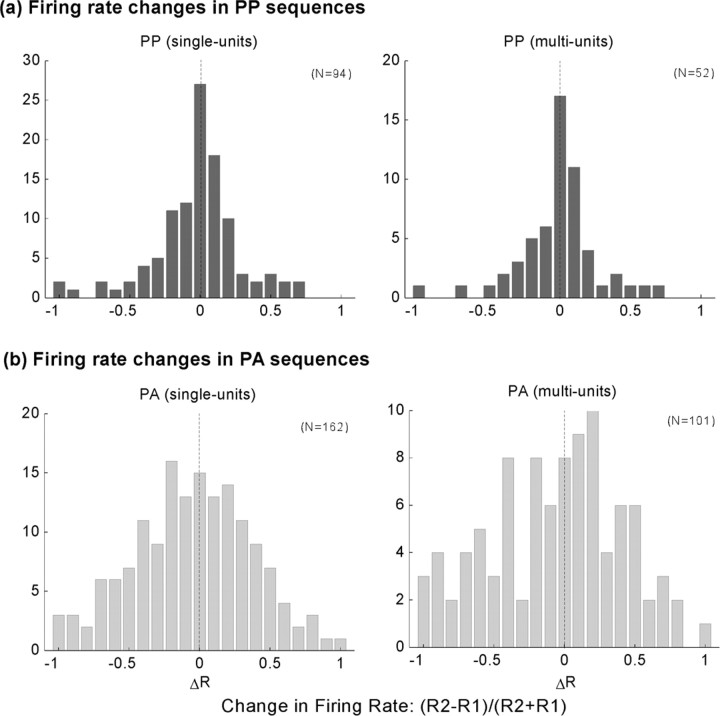
To form a reliable, consistent, and accurate representation of the acoustic scene, a reasonable conjecture is that cortical neurons maintain stable receptive fields after an early period of developmental plasticity. However, recent studies suggest that cortical neurons can be modified throughout adulthood and may change their response properties quite rapidly to reflect changing behavioral salience of certain sensory features. Because claims of adaptive receptive field plasticity could be confounded by intrinsic, labile properties of receptive fields themselves, we sought to gauge spontaneous changes in the responses of auditory cortical neurons. In the present study, we examined changes in a series of spectrotemporal receptive fields (STRFs) gathered from single neurons in successive recordings obtained over time scales of 30-120 min in primary auditory cortex (A1) in the quiescent, awake ferret. We used a global analysis of STRF shape based on a large database of A1 receptive fields. By clustering this STRF space in a data-driven manner, STRF sequences could be classified as stable or labile. We found that >73% of A1 neurons exhibited stable receptive field attributes over these time scales. In addition, we found that the extent of intrinsic variation in STRFs during the quiescent state was insignificant compared with behaviorally induced STRF changes observed during performance of spectral auditory tasks. Our results confirm that task-related changes induced by attentional focus on specific acoustic features were indeed confined to behaviorally salient acoustic cues and could be convincingly attributed to learning-induced plasticity when compared with "spontaneous" receptive field variability.
Figures
References
-
- Abut H. Vector quantization. Piscataway, NJ: IEEE; 1990.
-
- Aertsen A, Johannesma P. The spectro-temporal receptive field: a functional characteristic of auditory neurons. Biol Cybernetics. 1981;42:133–143. - PubMed
-
- Bair W. Visual receptive field organization. Curr Opin Neurobiol. 2005;15:459–464. - PubMed
-
- Breiman L, Friedman JH, Olshen RA, Stone CJ. Classification and regression trees. Boca Raton, FL: Chapman & Hall/CRC; 1984.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources