

Review
doi: 10.1039/b603600g.
Epub 2007 Jul 2.
The type I fatty acid and polyketide synthases: a tale of two megasynthases
Affiliations
- PMID: 17898897
- PMCID: PMC2263081
- DOI: 10.1039/b603600g
Item in Clipboard
Review
The type I fatty acid and polyketide synthases: a tale of two megasynthases
Nat Prod Rep.
2007 Oct.
Abstract
This review chronicles the synergistic growth of the fields of fatty acid and polyketide synthesis over the last century. In both animal fatty acid synthases and modular polyketide synthases, similar catalytic elements are covalently linked in the same order in megasynthases. Whereas in fatty acid synthases the basic elements of the design remain immutable, guaranteeing the faithful production of saturated fatty acids, in the modular polyketide synthases, the potential of the basic design has been exploited to the full for the elaboration of a wide range of secondary metabolites of extraordinary structural diversity.
Figures
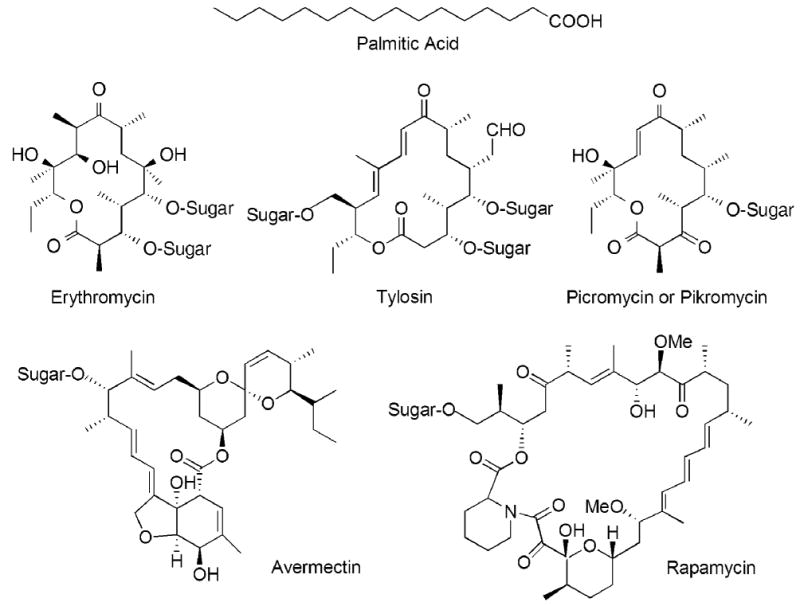
Chemical structures of some typical products of the animal FAS and modular PKSs. Many polyketides subsequently undergo modification by the addition of sugar residues.
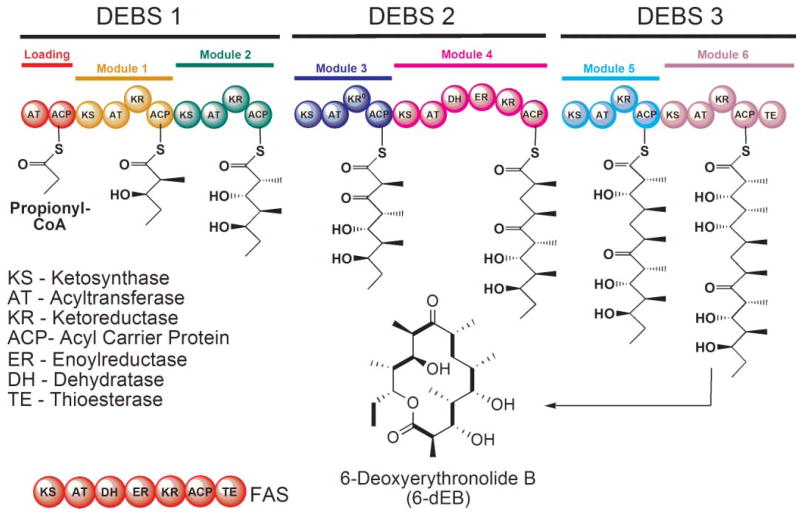
Domain organization of the animal FAS and the DEBS modular PKS. In the FAS, the AT domain is responsible for loading both the starter and chain-elongating substrates, whereas in modular PKSs, these substrates are loaded by separate ATs in the loading and chain-extending modules. The intermediates formed by each of the DEBS modules are shown together with the final 14-membered ring product. The KR domain in module 3 of DEBS is inactive.
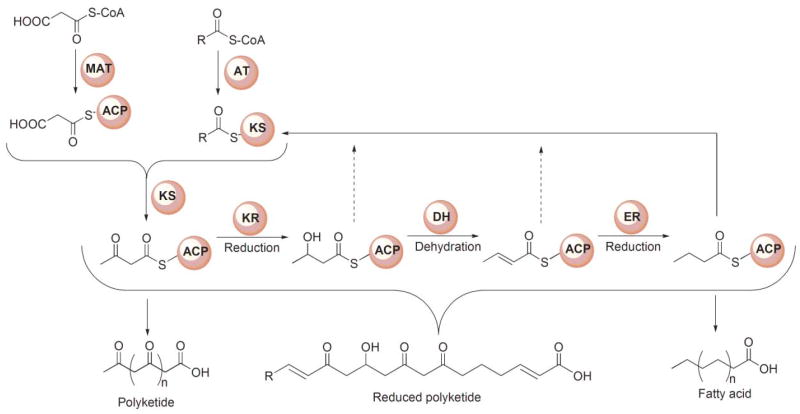
Generic reaction scheme for FASs and PKSs. The starter substrate for FASs typically is an acetyl moiety and the chain extender a malonyl moiety, whereas for PKSs, the starter is typically either acetyl or propionyl and the extender malonyl or methylmalonyl. In the FASs the β-ketoacyl moiety produced following each chain-extension reaction is completely reduced to a saturated carbon prior to the next condensation. In the modular PKSs, the β-ketoacyl moiety may undergo complete or partial reduction, leading to the formation of products that retain keto, hydroxyl or enoyl groups along the acyl chain.
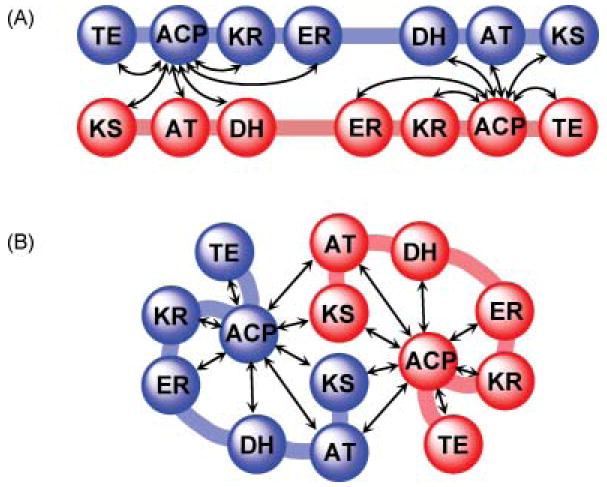
Cartoon representations of alternative models for the FAS. (A) The fully extended, head-to-tail orientated subunit model. (B) The head-to–head, coiled subunit model. In both models, arrows indicate the functional interactions occurring between domains, inter- and intrasubunit.
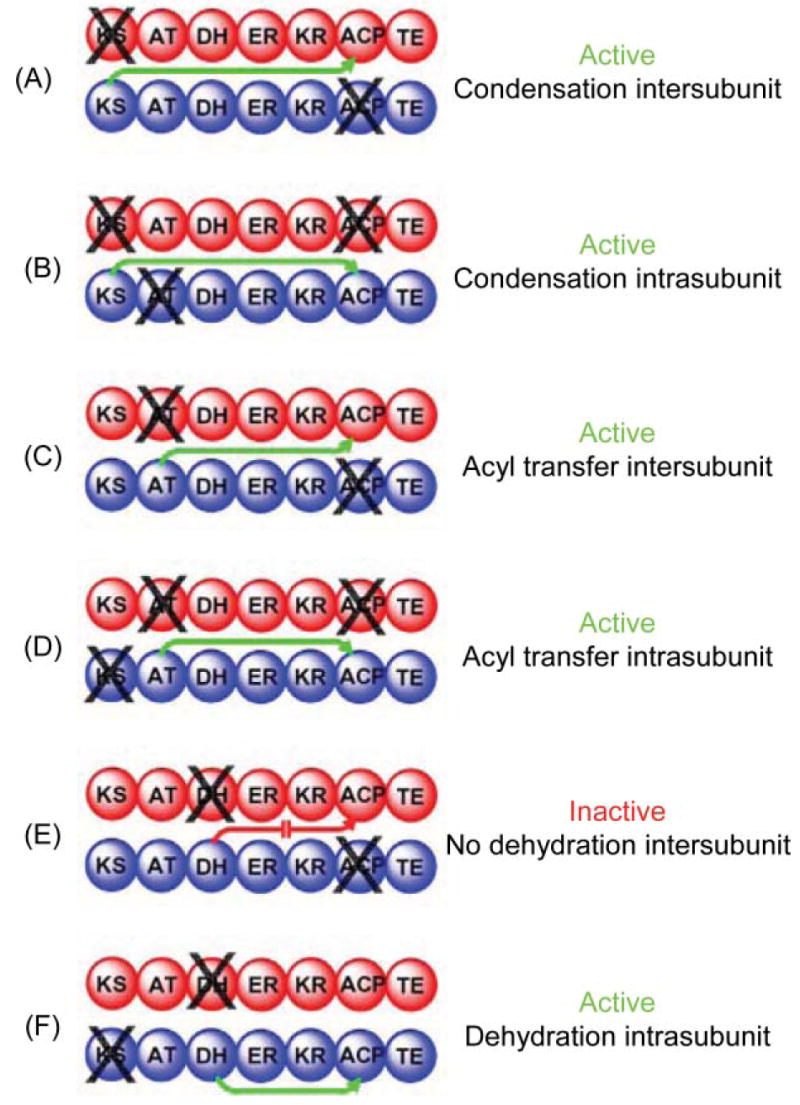
Mapping domain interactions in the FAS by mutant complementation analysis. The two individually mutated FAS subunits forming each heterodimer are colored red and blue and the site of the knockout mutation is indicated by X. Green arrows indicate which catalytic domains engage in functional interactions with an ACP domain. A red arrow indicates the blocked interaction responsible for an inactive heterodimer.
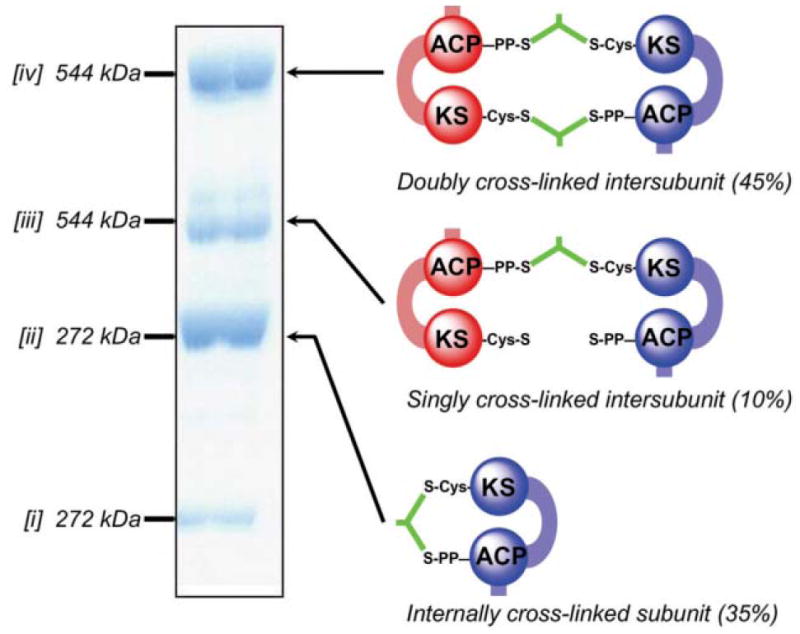
Cross-linking of FAS by dibromopropanone. The SDS–poly-acrylamide gel shows the three new cross-linked species [ii–iv] formed by treatment of the wild-type FAS with dibromopropanone. Species [i] represent residual subunits that have not undergone cross-linking. Molecular masses were determined by mass spectrometry. The identities of the cross-linked species were determined by performing cross-linking on a panel of mutants lacking either the ACP 4-phosphopantetheine or the KS active site cysteine on one or both subunits. The cartoons on the right illustrate the mode of cross-linking, either inter- or intra subunit.
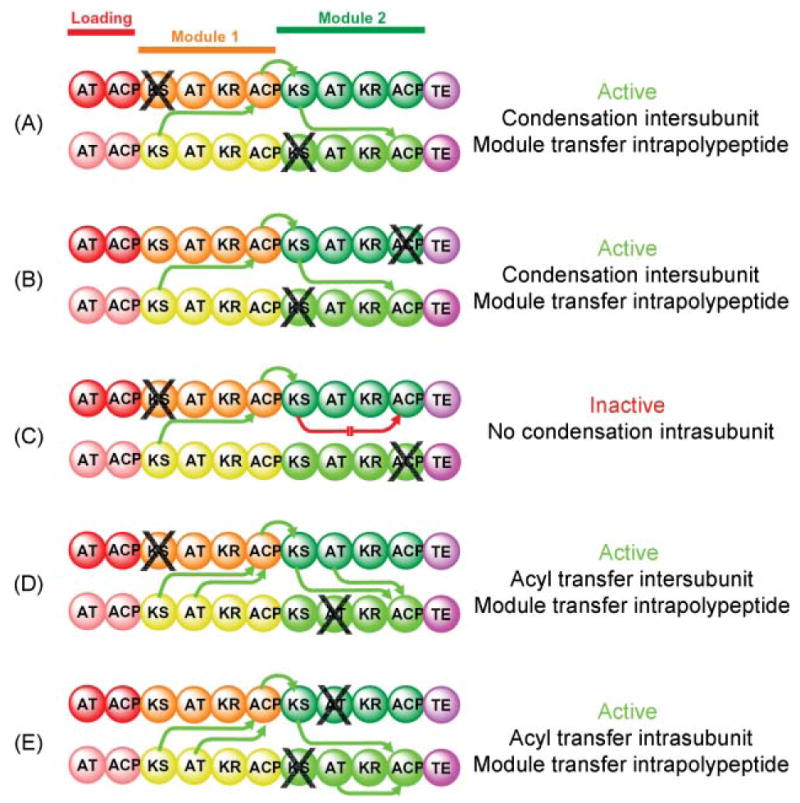
Mapping domain interactions in the DEBS PKS by mutant complementation analysis. The subunits used in this study were constructed by fusing the TE domain from module 6 to the C-terminus of the ACP domain in module 2. The site of a knockout mutation is indicated by X. The parental mutant homodimers were mixed together, the subunits were randomized to give a preparation consisting of hetero-and homodimers, then assayed for activity. Green arrows indicate which catalytic domains engage in functional interactions with an ACP domain and the directionality of intermodular substrate transfer. A red arrow indicates the blocked interaction responsible for an inactive heterodimer.
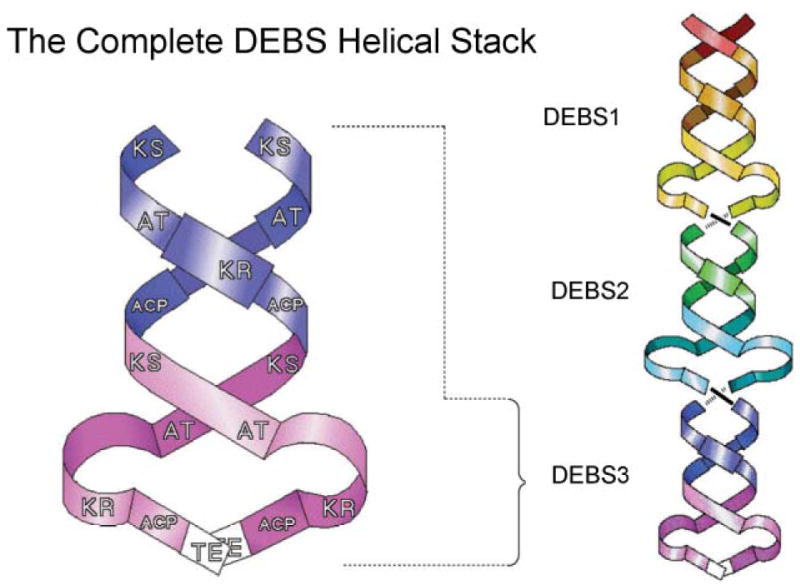
The Cambridge double helix model for modular PKSs. The core of the interwound helical dimer is formed by the KS, AT and ACP domains and the optional β-carbon processing enzymes loop away from the helix. Reproduced from ref. with permission.
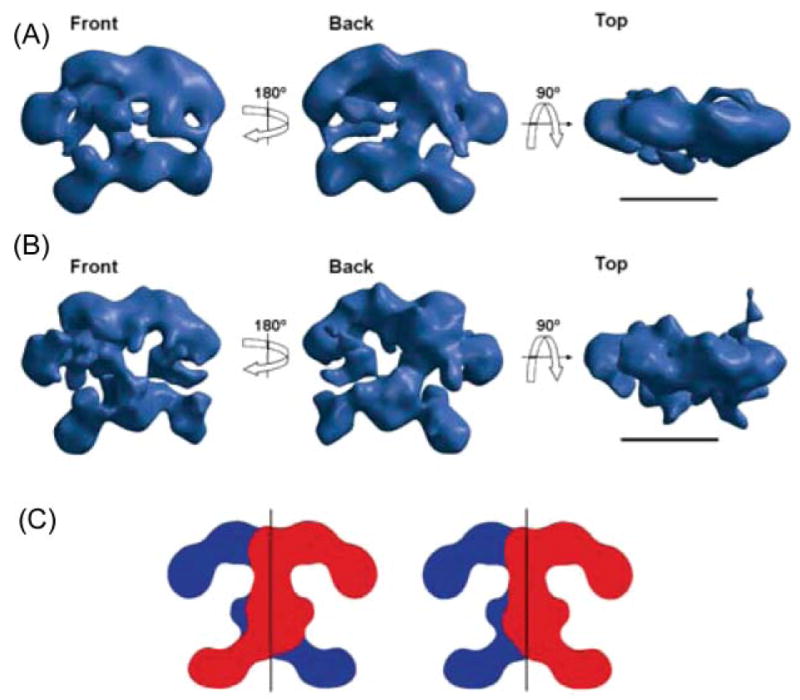
Structure of FAS determined by electron microscopy. (A) Calculated using the random conical tilt method from images of single particles of the Cys161Gln rat FAS mutant imaged under turnover conditions and preserved in negative stain; resolution ~30 Å. (B) Reconstruction of the same mutant, also imaged under turnover conditions, but preserved in amorphous ice; resolution ~16 Å. Scale bar 100 Å. (C) Alternative arrangements of the two subunits that are consistent with the structure of the dimer. Reproduced with permission from ref. .
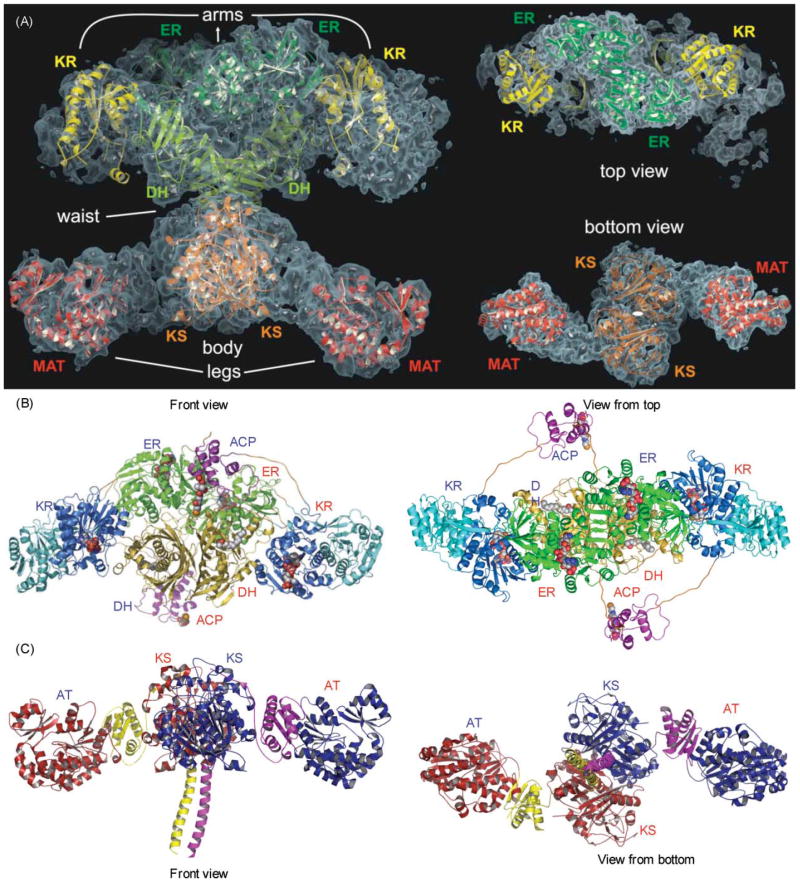
Crystallographic analysis of the megasynthases. (A) Structural overview of the porcine FAS. Fitted homologous domains are shown with a semitransparent surface representation of the experimental electron density. Left, front view; right, top and bottom views. Reproduced with permission from ref. . (B) Modeled structure of the section of a hypothetical PKS containing DH, ER, KR and ACP domains. Polyglycine linkers, in orange, were built to connect the domains. The molecule bound to DH is N,N-dimethyldodecylamineoxide (detergent bound in the TEII template structure) and to the reductases, NADPH. The phosphopantetheine of ACP is also displayed. Left, front view; right, top view. Reproduced with permission from ref. . (C) X-Ray structure of the KS–AT didomain from DEBS module 5, The three linker regions of the DEBS KS–AT include the N-terminal helices, the KS–AT linker and the C-terminal linker, colored in yellow in monomer A and purple in monomer B. In the porcine FAS model, these three linker regions contain electron density, but models of these regions were not built. Left, front view; right, bottom view. Adapted from ref. .
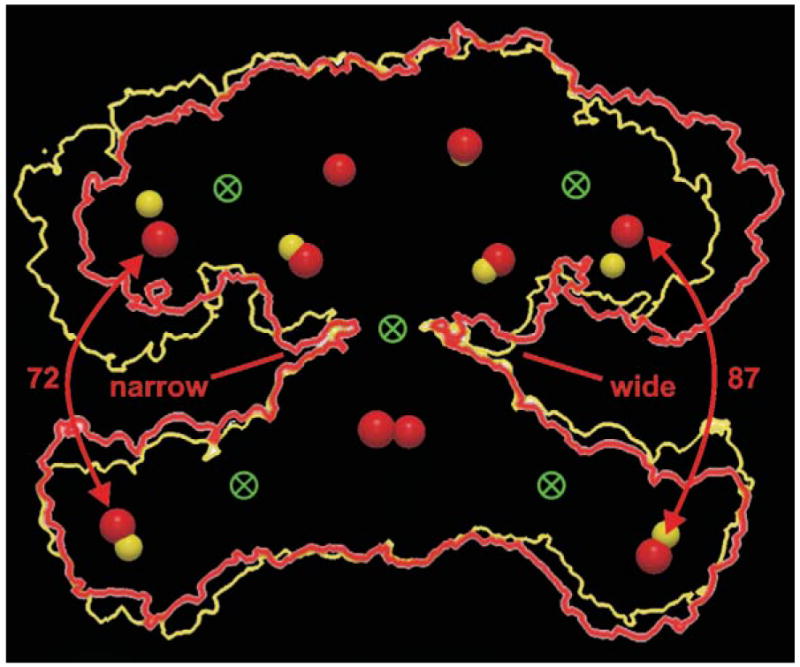
Interdomain hinges and conformational flexibility. An outline of the FAS dimer structure (red) was rotated 180° and the resulting outline (yellow) superimposed on the original. The active sites of the two structures are shown as red and yellow spheres and the possible location of hinges that could facilitate the conformational changes required to produce the two structures are shown as crossed circles. Reproduced with permission from ref. .
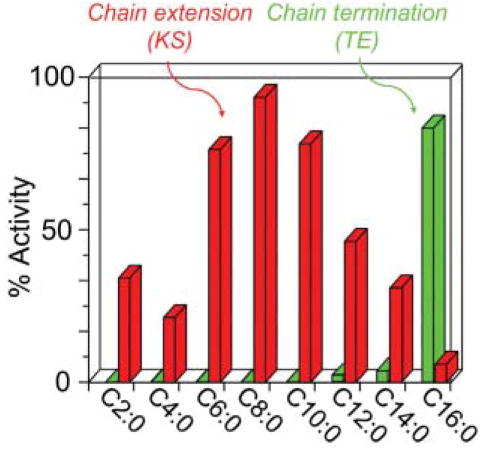
Substrate specificities of the chain-extending and chain-termination enzymes of the animal FAS. Chain-extension activity represents the activity of the KS domain interthiol acyltransferase and chain termination activity, the activity of the TE domain, both assayed using acyl-CoAs as model substrates.
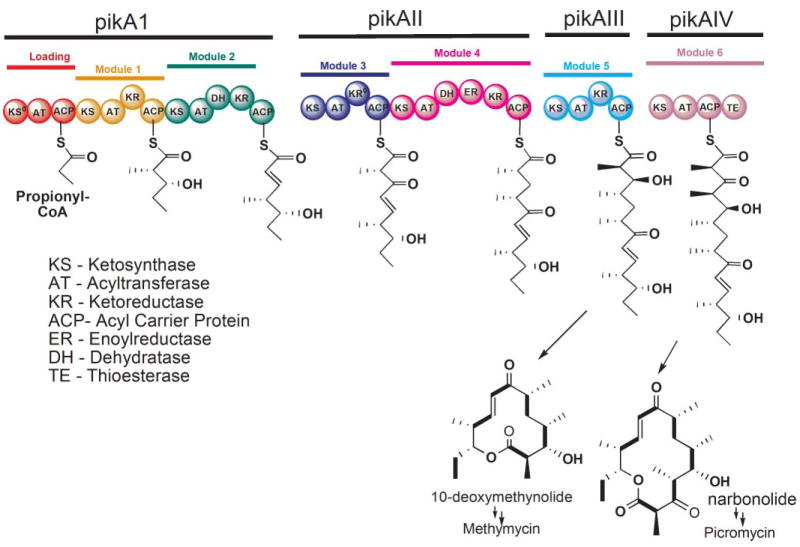
The modular architecture of the ‘Pik’ PKS. An acyl chain released from module 5 and cyclized forms 10-deoxymethynolide, whereas cyclization of the product released from module 6 forms narbonolide.
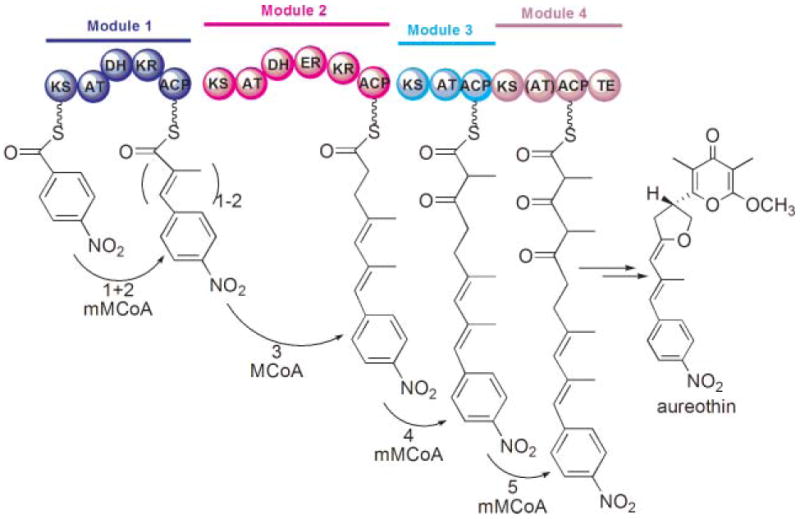
Programmed iteration by the aureothin modular PKS. An unusual starter moiety, p-nitrobenzoyl, is subjected to two rounds of chain extension in module 1 by methylmalonyl-CoA (mMCoA) followed by a normal chain-extension process successively in modules 2–4.
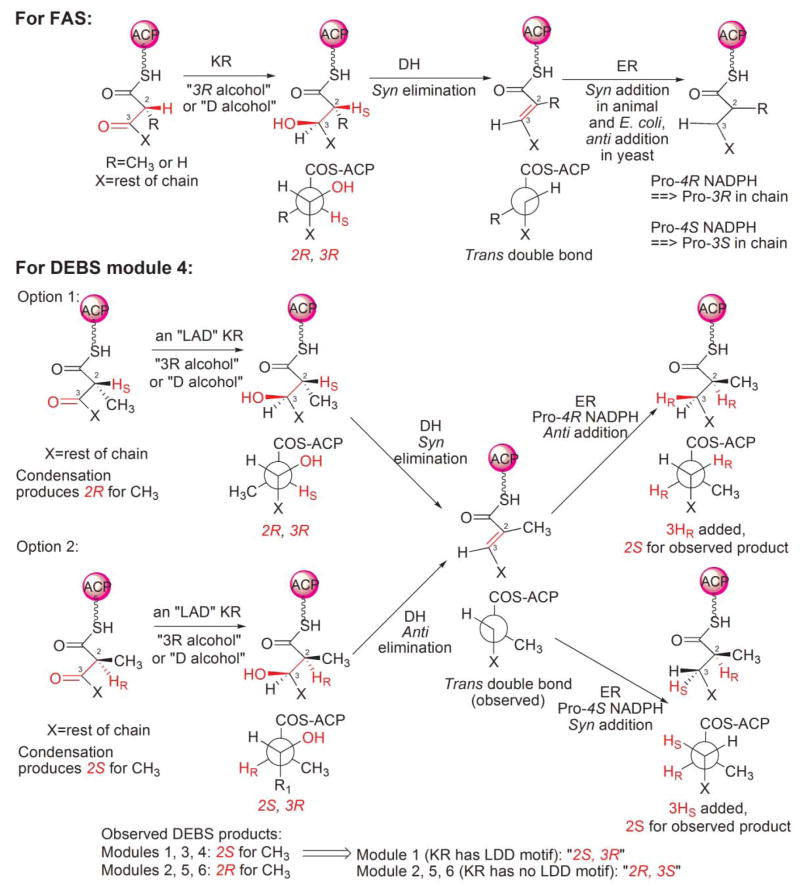
The stereochemistry of FAS and PKS.
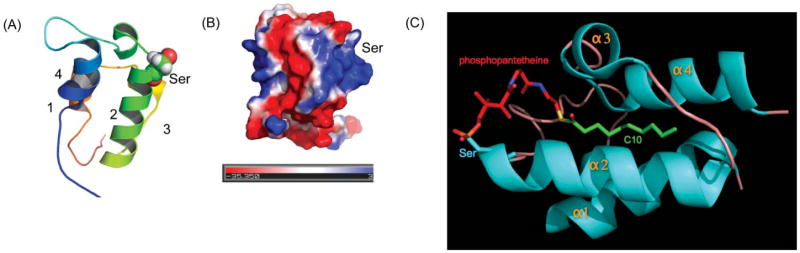
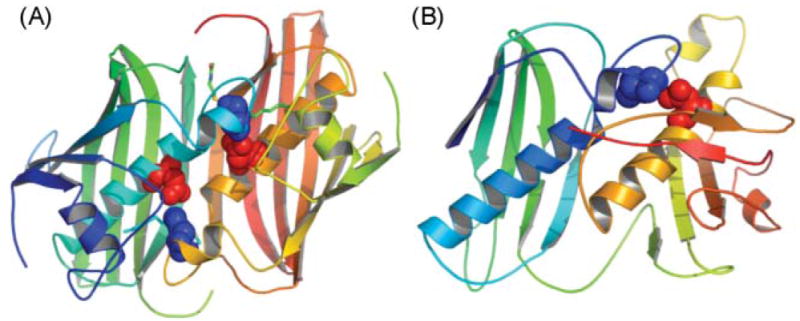
The ACP structure. (A) Ribbon diagram of the rat FAS ACP (2PNG), colored blue to red, corresponding to N to C termini., The post-translationally modified serine residue on helix 2 is shown. (B) The ACP surface electrostatic potential colored from red (negative) to blue (positive). (C) The crystal structure of the decanoyl thioester of E. coli ACP. The phosphopantetheine moiety is shown in red and the decanoyl moiety in green. Adapted from ref. .
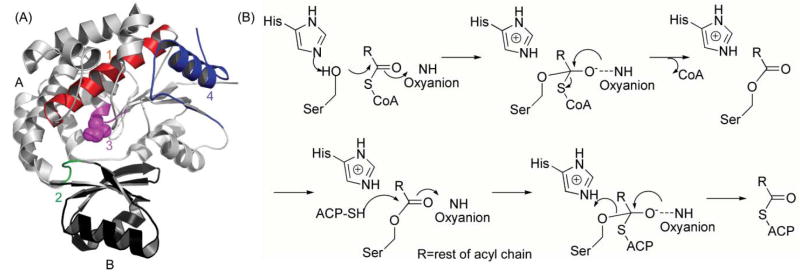
(A) The DEBS AT5 domain, showing the overall fold with the active site Ser in purple, and the substrate-binding pocket under the Ser in the cleft formed by domains A and B. The four structural elements discussed in the text (items 1–4) that are important for specificity are highlighted in red, purple, green and blue, respectively. (B) Proposed catalytic mechanism of the AT domain. R = rest of acyl chain.
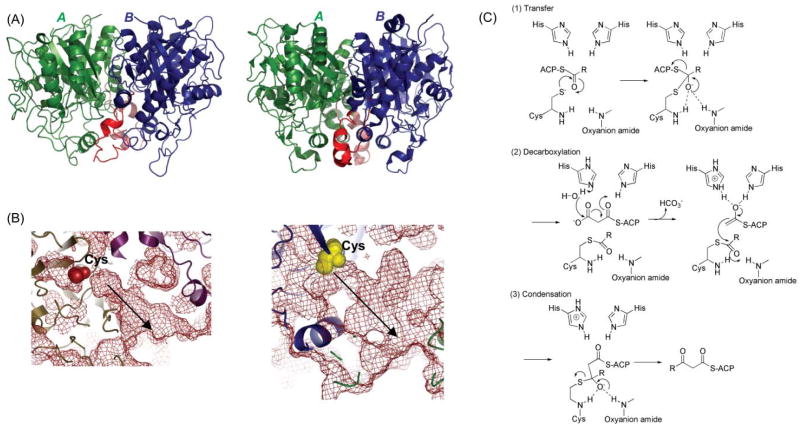
The KS domain. (A) Overall fold comparison: left, DEBS KS; right, FAS KS; monomers are shown in green and blue and the dimer interface helix-loop in red. (B) Molecular basis of substrate specificity. The FAS KS channel is narrower with limited flexibility, reflecting its limited substrate tolerance for branched acyl chains, while the DEBS KS5 channel is much wider, reflecting its substrate tolerance; the substrate-binding pockets are identified by arrows. The active site Cys is shown as yellow or red spheres in both panels. (C) Proposed mechanism.
The KR domain. Crystal structure of the DEBS1 KR domain. The catalytic subdomain is shown in blue, the structural subdomain in red, the active site residues in purple, the phenylalanine, proline and glycine residues important for substrate binding in orange, the ‘LDD’ loop in yellow and NADP in green. Adapted from ref. .
The DH domain. The E. coli DH, FabA (A) and modeled DH domain of FAS (B) exhibit two hotdog folds. The structures are colored from blue to red, from N- to C-terminus. The active site histidine (blue) and aspartate (red) residues are shown.
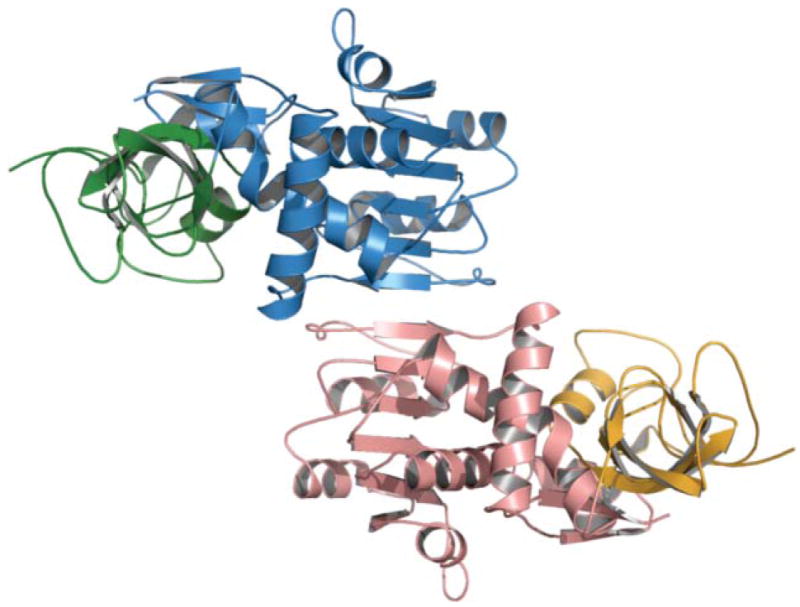
The rat FAS ER homology model (built using the C-alpha trace of the porcine FAS model) shows a tightly interacting dimer. The nucleotide-binding and catalytic domains are colored blue and green in monomer A and red and orange in monomer B. The dimer interface spans 10 β-strands located in the nucleotide-binding domain.
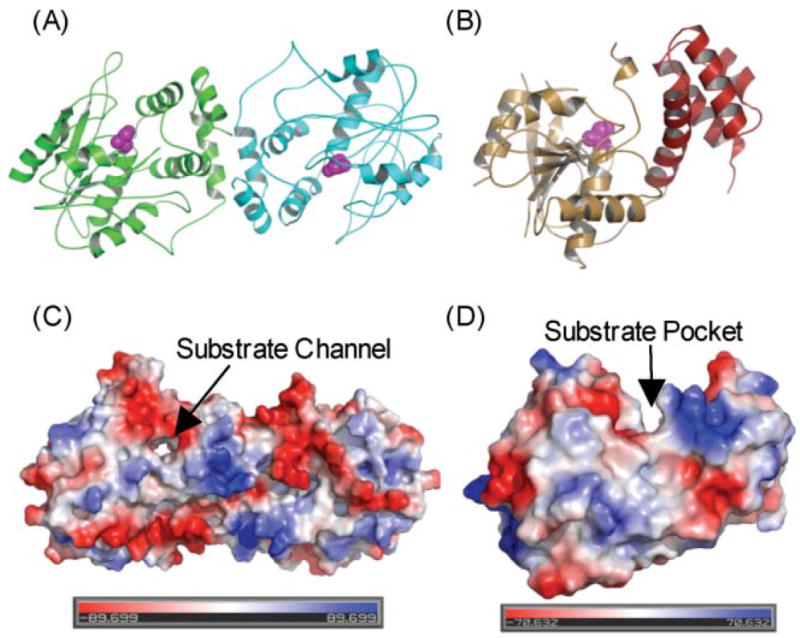
The TE domain. (A) Overall fold of DEBS TE dimer; monomer A in green and monomer B in blue. (B) Overall fold of FAS TE monomer; domain A in orange, domain B in red. The active-site serine is shown in purple. (C) Molecular surface of DEBS TE and FAS TE, colored in red (negative) and blue (positive) to reflect electrostatic potential.
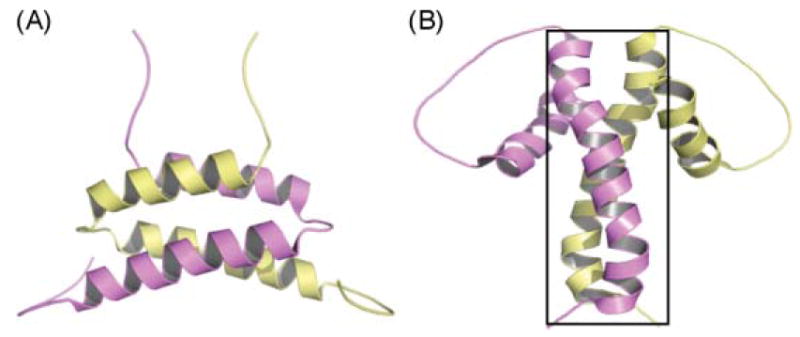
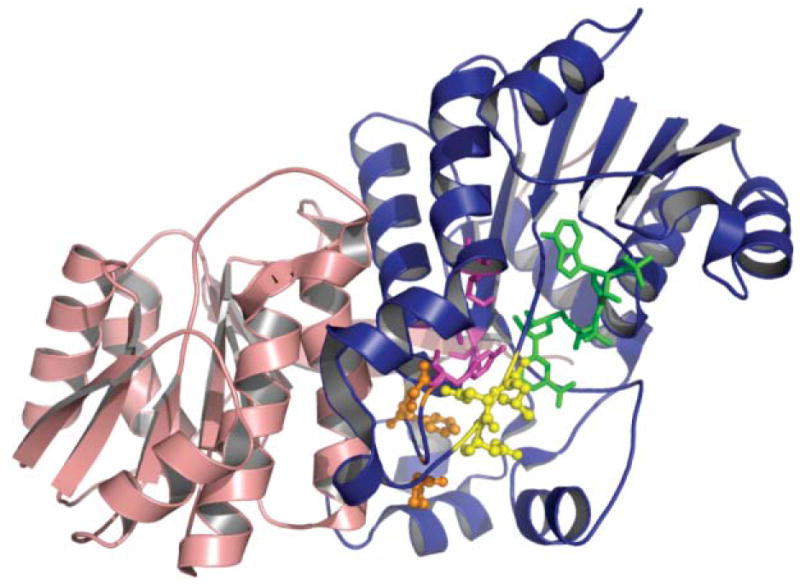
The A and B domains of Dock 2–3. (A) Domain A, (B) domain B. Monomer A is colored in yellow and monomer B in purple. The N-terminal coiled-coil observed in the crystal structure of DEBS module 5 KS–AT is enclosed in a box.
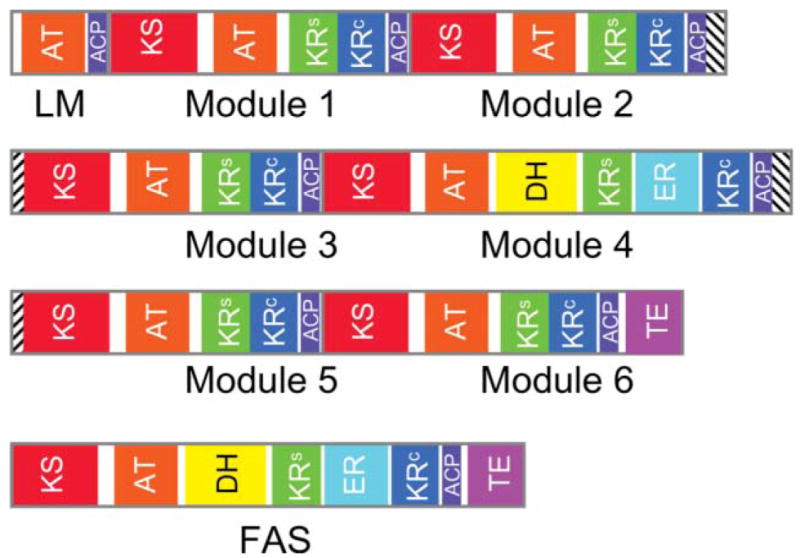
Revised domain maps for the animal FAS and a modular PKS. The domain maps for the loading module (LM) and six extender modules of DEBS include the interpolypeptide docking domains (cross-hatched). The KR domain of module 3 is inactive. In both the FAS and modular PKS, the region previously referred to as the central core is assigned to the KR structural subdomain (KRs) and, where present, the second subdomain of the DH. Thus the KR catalytic (KRc) and structural subdomains are contiguous in DEBS modules 1, 2, 3, 5 and 6, but are split by the ER domain in module 4 and in the FAS.
References
-
- Staunton J, Weissman KJ. Nat Prod Rep. 2001;18:380. - PubMed
-
- Shen B. Curr Opin Chem Biol. 2003;7:285. - PubMed
-
- Taylor WC. Snows of yesteryear: J Norman Collie, mountaineer. Holt, Rinehart and Winston; Toronto: 1973.
-
- Collie J. J Chem Soc, Trans. 1907;91:1806.
-
- Raper H. J Chem Soc, Trans. 1907;91:1831.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources