

Disease-associated mutant alpha-actinin-4 reveals a mechanism for regulating its F-actin-binding affinity
- PMID: 17901210
- PMCID: PMC2042165
- DOI: 10.1073/pnas.0702451104
Disease-associated mutant alpha-actinin-4 reveals a mechanism for regulating its F-actin-binding affinity
Abstract
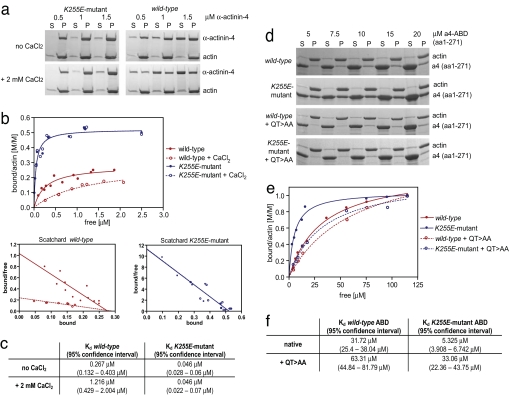
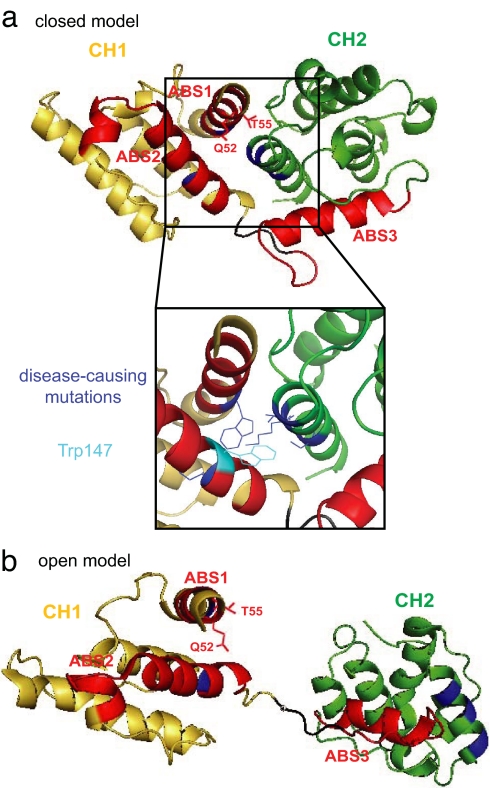
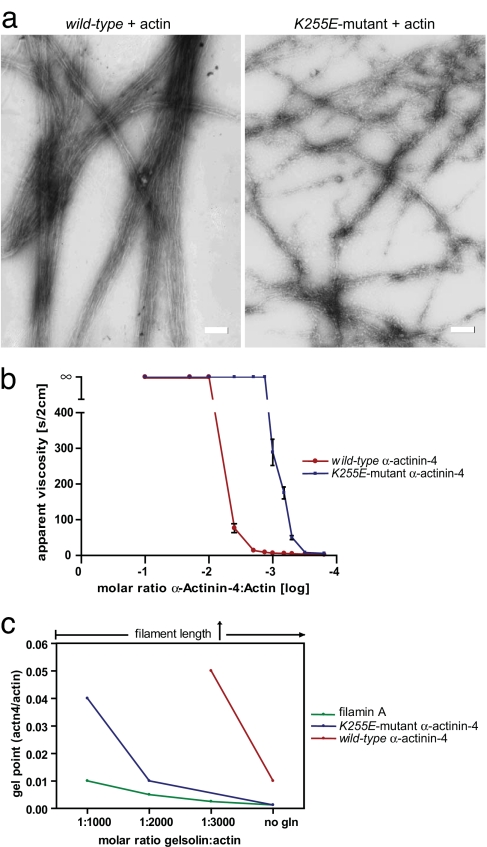
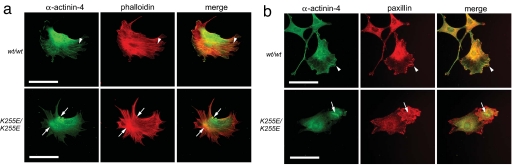
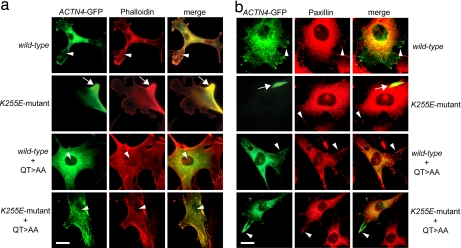
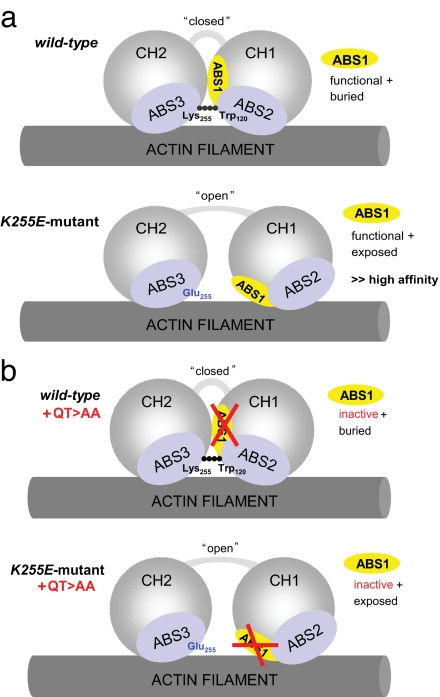
Alpha-actinin-4 is a widely expressed protein that employs an actin-binding site with two calponin homology domains to crosslink actin filaments (F-actin) in a Ca(2+)-sensitive manner in vitro. An inherited, late-onset form of kidney failure is caused by point mutations in the alpha-actinin-4 actin-binding domain. Here we show that alpha-actinin-4/F-actin aggregates, observed in vivo in podocytes of humans and mice with disease, likely form as a direct result of the increased actin-binding affinity of the protein. We document that exposure of a buried actin-binding site 1 in mutant alpha-actinin-4 causes an increase in its actin-binding affinity, abolishes its Ca(2+) regulation in vitro, and diverts its normal localization from actin stress fibers and focal adhesions in vivo. Inactivation of this buried actin-binding site returns the affinity of the mutant to that of the WT protein and abolishes aggregate formation in cells. In vitro, actin filaments crosslinked by the mutant alpha-actinin-4 exhibit profound changes of structural and biomechanical properties compared with WT alpha-actinin-4. On a molecular level, our findings elucidate the physiological importance of a dynamic interaction of alpha-actinin with F-actin in podocytes in vivo. We propose that a conformational change with full exposure of actin-binding site 1 could function as a switch mechanism to regulate the actin-binding affinity of alpha-actinin and possibly other calponin homology domain proteins under physiological conditions.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Crystal structure of the actin-binding domain of alpha-actinin-4 Lys255Glu mutant implicated in focal segmental glomerulosclerosis.J Mol Biol. 2008 Feb 15;376(2):317-24. doi: 10.1016/j.jmb.2007.11.084. Epub 2007 Dec 4. J Mol Biol. 2008. PMID: 18164029 Free PMC article.
-
Congenital macrothrombocytopenia-linked mutations in the actin-binding domain of α-actinin-1 enhance F-actin association.FEBS Lett. 2016 Mar;590(6):685-95. doi: 10.1002/1873-3468.12101. Epub 2016 Mar 1. FEBS Lett. 2016. PMID: 26879394
-
Dynamic Regulation of α-Actinin's Calponin Homology Domains on F-Actin.Biophys J. 2016 Mar 29;110(6):1444-55. doi: 10.1016/j.bpj.2016.02.024. Biophys J. 2016. PMID: 27028653 Free PMC article.
-
Molecular biology of actin binding proteins: evidence for a common structural domain in the F-actin binding sites of gelsolin and alpha-actinin.J Cell Sci Suppl. 1991;14:91-4. doi: 10.1242/jcs.1991.supplement_14.19. J Cell Sci Suppl. 1991. PMID: 1653252 Review.
-
Structure and evolution of the actin crosslinking proteins.Bioessays. 1991 May;13(5):219-26. doi: 10.1002/bies.950130504. Bioessays. 1991. PMID: 1892474 Review.
Cited by
-
Phylogenetic analysis of gene structure and alternative splicing in alpha-actinins.Mol Biol Evol. 2010 Apr;27(4):773-80. doi: 10.1093/molbev/msp268. Epub 2009 Nov 6. Mol Biol Evol. 2010. PMID: 19897525 Free PMC article.
-
Alpha-actinin-4 and CLP36 protein deficiencies contribute to podocyte defects in multiple human glomerulopathies.J Biol Chem. 2011 Sep 2;286(35):30795-30805. doi: 10.1074/jbc.M111.255984. Epub 2011 Jun 16. J Biol Chem. 2011. PMID: 21680739 Free PMC article.
-
MTBP suppresses cell migration and filopodia formation by inhibiting ACTN4.Oncogene. 2013 Jan 24;32(4):462-70. doi: 10.1038/onc.2012.69. Epub 2012 Feb 27. Oncogene. 2013. PMID: 22370640 Free PMC article.
-
Cell-matrix adhesion of podocytes in physiology and disease.Nat Rev Nephrol. 2013 Apr;9(4):200-10. doi: 10.1038/nrneph.2012.291. Epub 2013 Jan 22. Nat Rev Nephrol. 2013. PMID: 23338211 Review.
-
Dynamic actin cross-linking governs the cytoplasm's transition to fluid-like behavior.Mol Biol Cell. 2020 Jul 21;31(16):1744-1752. doi: 10.1091/mbc.E19-09-0504. Epub 2020 Jun 24. Mol Biol Cell. 2020. PMID: 32579489 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
