

The requirement for Phr1 in CNS axon tract formation reveals the corticostriatal boundary as a choice point for cortical axons
- PMID: 17901218
- PMCID: PMC2000324
- DOI: 10.1101/gad.1592107
The requirement for Phr1 in CNS axon tract formation reveals the corticostriatal boundary as a choice point for cortical axons
Abstract
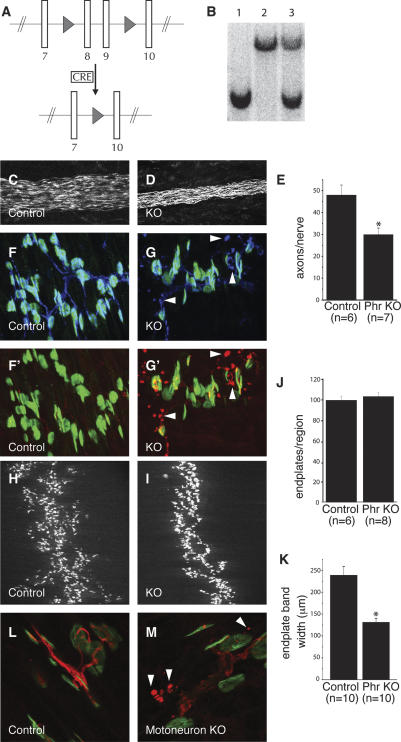
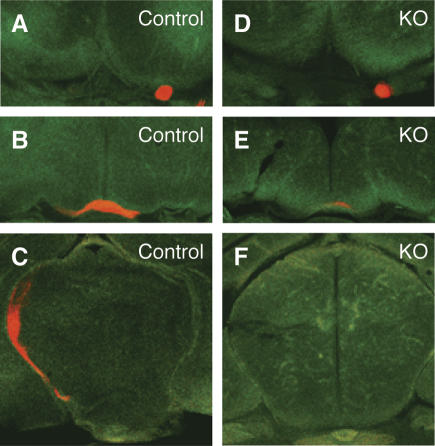
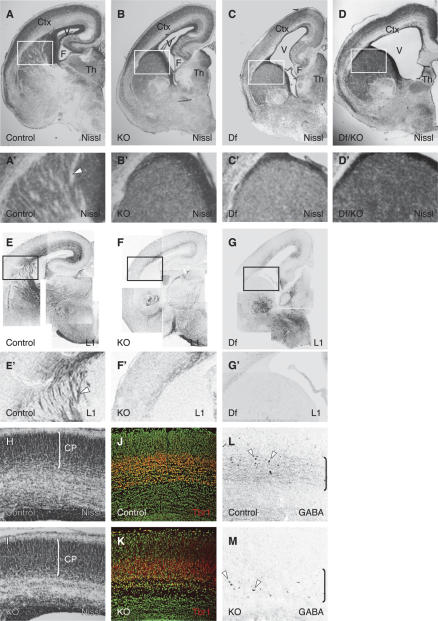
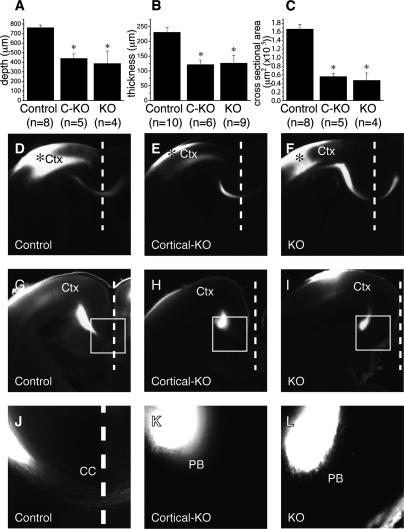
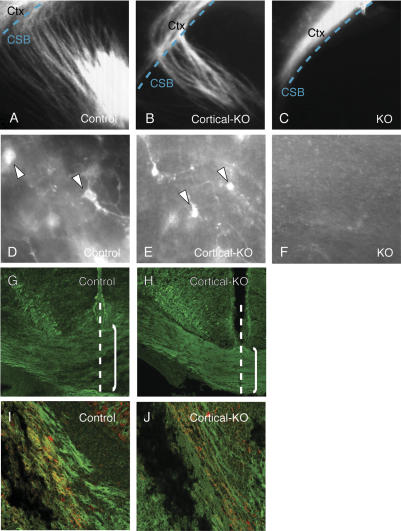
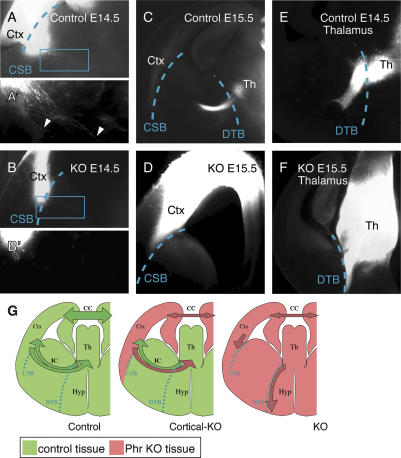
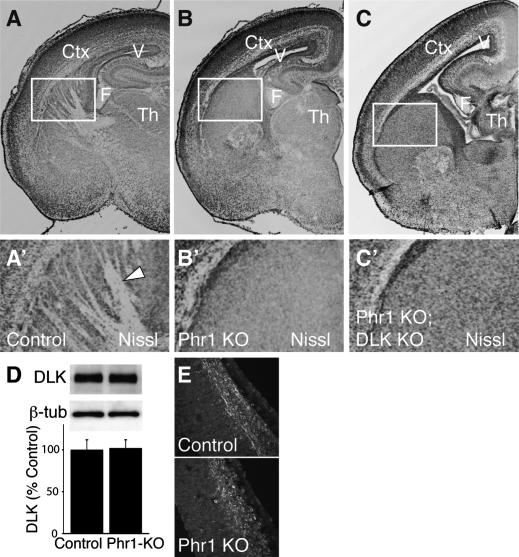
Phr1 is the single well-conserved murine ortholog of the invertebrate ubiquitin ligase genes highwire (in Drosophila) and rpm-1 (in Caenorhabditis elegans). The function and mechanism of action of highwire and rpm-1 are similar--both cell-autonomously regulate synaptogenesis by down-regulating the ortholog of the mitogen-activated protein kinase kinase kinase dual leucine zipper kinase (MAPKKK DLK). Here, using a targeted conditional mutant, we demonstrate that Phr1 also plays essential roles in mammalian neural development. As in invertebrates, Phr1 functions cell-autonomously to sculpt motor nerve terminals. In addition, Phr1 plays essential roles in the formation of major CNS axon tracts including those of the internal capsule, in part via cell-nonautonomous mechanisms, and these results reveal a choice point for cortical axons at the corticostriatal boundary. Furthermore, whereas the neurite morphology phenotypes of highwire and rpm-1 are suppressed by loss of DLK in flies and worms, Phr1-dependent CNS defects persist in Phr1, DLK double mutants. Thus, in the mammalian nervous system Phr1 is required for formation of major CNS axon tracts via a mechanism that is both cell-nonautonomous and independent of DLK.
Figures
References
-
- Bagri A., Marin O., Plump A.S., Mak J., Pleasure S.J., Rubenstein J.L., Tessier-Lavigne M., Marin O., Plump A.S., Mak J., Pleasure S.J., Rubenstein J.L., Tessier-Lavigne M., Plump A.S., Mak J., Pleasure S.J., Rubenstein J.L., Tessier-Lavigne M., Mak J., Pleasure S.J., Rubenstein J.L., Tessier-Lavigne M., Pleasure S.J., Rubenstein J.L., Tessier-Lavigne M., Rubenstein J.L., Tessier-Lavigne M., Tessier-Lavigne M. Slit proteins prevent midline crossing and determine the dorsoventral position of major axonal pathways in the mammalian forebrain. Neuron. 2002;33:233–248. - PubMed
-
- Bareyre F.M., Kerschensteiner M., Misgeld T., Sanes J.R., Kerschensteiner M., Misgeld T., Sanes J.R., Misgeld T., Sanes J.R., Sanes J.R. Transgenic labeling of the corticospinal tract for monitoring axonal responses to spinal cord injury. Nat. Med. 2005;11:1355–1360. - PubMed
-
- Burgess R.W., Peterson K.A., Johnson M.J., Roix J.J., Welsh I.C., O’Brien T.P., Peterson K.A., Johnson M.J., Roix J.J., Welsh I.C., O’Brien T.P., Johnson M.J., Roix J.J., Welsh I.C., O’Brien T.P., Roix J.J., Welsh I.C., O’Brien T.P., Welsh I.C., O’Brien T.P., O’Brien T.P. Evidence for a conserved function in synapse formation reveals Phr11 as a candidate gene for respiratory failure in newborn mice. Mol. Cell. Biol. 2004;24:1096–1105. - PMC - PubMed
-
- Carney R.S., Alfonso T.B., Cohen D., Dai H., Nery S., Stocia B., Slotkin J., Bregman B.S., Fishell G., Corbin J.G., Alfonso T.B., Cohen D., Dai H., Nery S., Stocia B., Slotkin J., Bregman B.S., Fishell G., Corbin J.G., Cohen D., Dai H., Nery S., Stocia B., Slotkin J., Bregman B.S., Fishell G., Corbin J.G., Dai H., Nery S., Stocia B., Slotkin J., Bregman B.S., Fishell G., Corbin J.G., Nery S., Stocia B., Slotkin J., Bregman B.S., Fishell G., Corbin J.G., Stocia B., Slotkin J., Bregman B.S., Fishell G., Corbin J.G., Slotkin J., Bregman B.S., Fishell G., Corbin J.G., Bregman B.S., Fishell G., Corbin J.G., Fishell G., Corbin J.G., Corbin J.G. Cell migration along the lateral cortical stream to the developing basal telencephalic limbic system. J. Neurosci. 2006;26:11562–11574. - PMC - PubMed
-
- Chapouton P., Schuurmans C., Guillemot F., Gotz M., Schuurmans C., Guillemot F., Gotz M., Guillemot F., Gotz M., Gotz M. The transcription factor neurogenin 2 restricts cell migration from the cortex to the striatum. Development. 2001;128:5149–5159. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials