

Platelet-VWF complexes are preferred substrates of ADAMTS13 under fluid shear stress
- PMID: 17901248
- PMCID: PMC2200832
- DOI: 10.1182/blood-2007-05-093021
Platelet-VWF complexes are preferred substrates of ADAMTS13 under fluid shear stress
Abstract
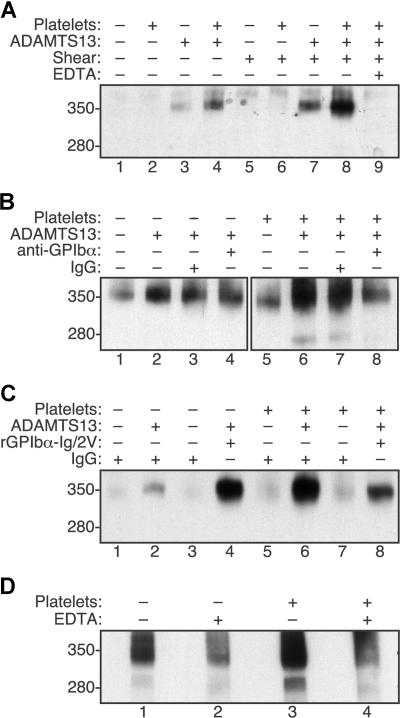
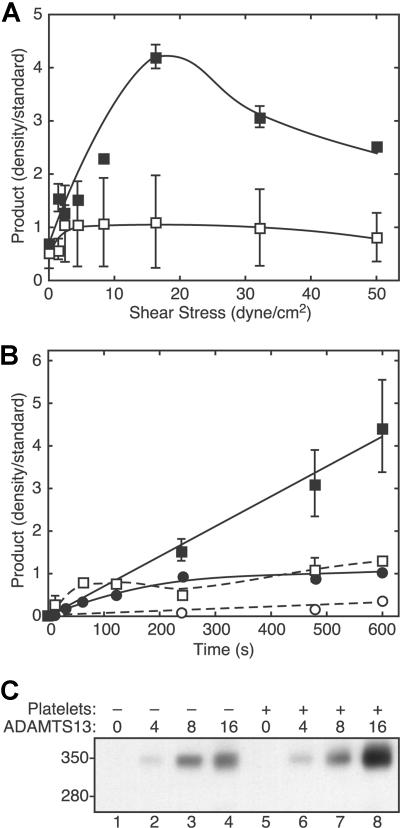
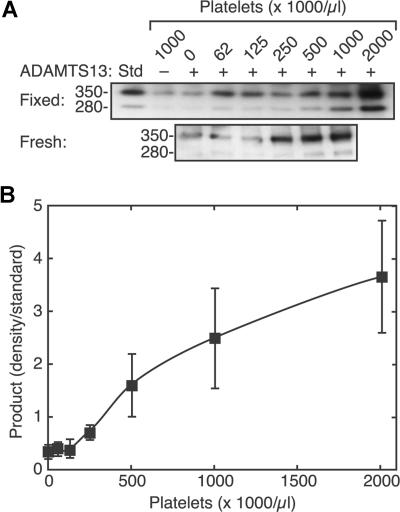
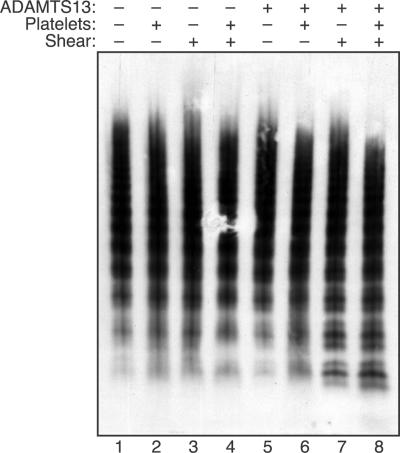
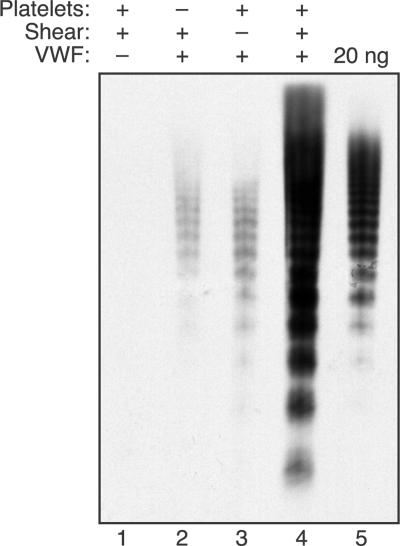
Endothelial cells secrete prothrombotic ultralarge von Willebrand factor (VWF) multimers, and the metalloprotease ADAMTS13 cleaves them into smaller, less dangerous multimers. This reaction is stimulated by tensile force applied to the VWF substrate, which may occur on cell surfaces or in the circulating blood. The cleavage of soluble VWF by ADAMTS13 was accelerated dramatically by a combination of platelets and fluid shear stress applied in a cone-plate viscometer. Platelet-dependent cleavage of VWF was blocked by an anti-GPIbalpha monoclonal antibody or by a recombinant soluble fragment of GPIbalpha that prevents platelet-VWF binding. Multimeric gel analysis showed that shear and platelet-dependent cleavage consumed large VWF multimers. Therefore, ADAMTS13 preferentially acts on platelet-VWF complexes under fluid shear stress. This reaction is likely to account for a majority of VWF proteolysis after secretion and to determine the steady-state size distribution of circulating VWF multimers in vivo.
Figures
References
-
- Sadler JE. Biochemistry and genetics of von Willebrand factor. Annu Rev Biochem. 1998;67:395–424. - PubMed
-
- Ruggeri ZM. Von Willebrand factor, platelets and endothelial cell interactions. J Thromb Haemost. 2003;1:1335–1342. - PubMed
-
- Zheng ×, Chung D, Takayama TK, Majerus EM, Sadler JE, Fujikawa K. Structure of von Willebrand factor-cleaving protease (ADAMTS13), a metalloprotease involved in thrombotic thrombocytopenic purpura. J Biol Chem. 2001;276:41059–41063. - PubMed
-
- Levy GG, Nichols WC, Lian EC, et al. Mutations in a member of the ADAMTS gene family cause thrombotic thrombocytopenic purpura. Nature. 2001;413:488–494. - PubMed
-
- Soejima K, Mimura N, Hirashima M, et al. A novel human metalloprotease synthesized in the liver and secreted into the blood: possibly, the von Willebrand factor-cleaving protease? J Biochem. 2001;130:475–480. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
