

Factors that regulate embryonic gustatory development
- PMID: 17903280
- PMCID: PMC1995452
- DOI: 10.1186/1471-2202-8-S3-S4
Factors that regulate embryonic gustatory development
Abstract
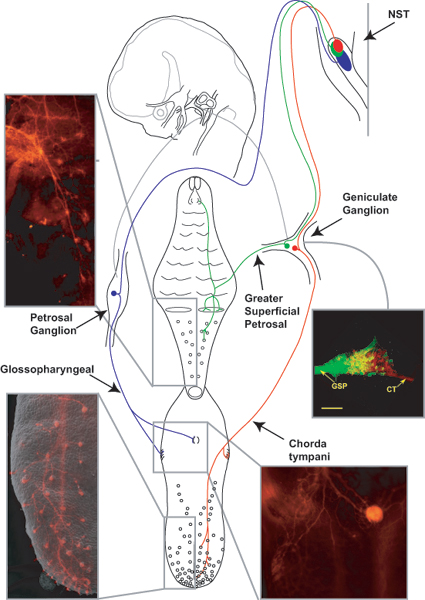
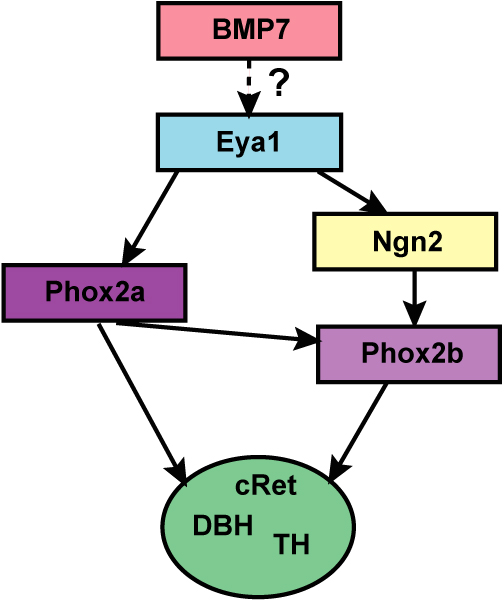
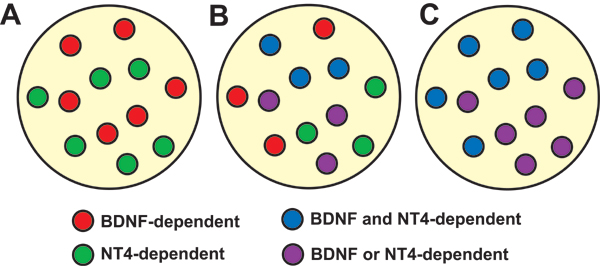
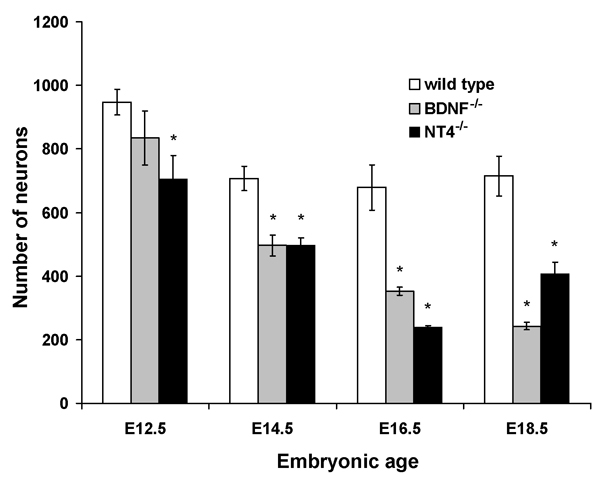
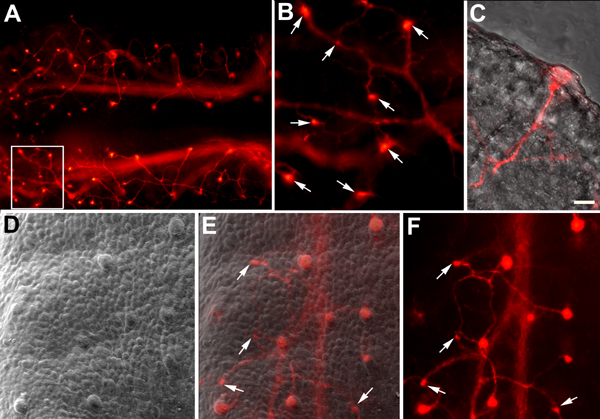
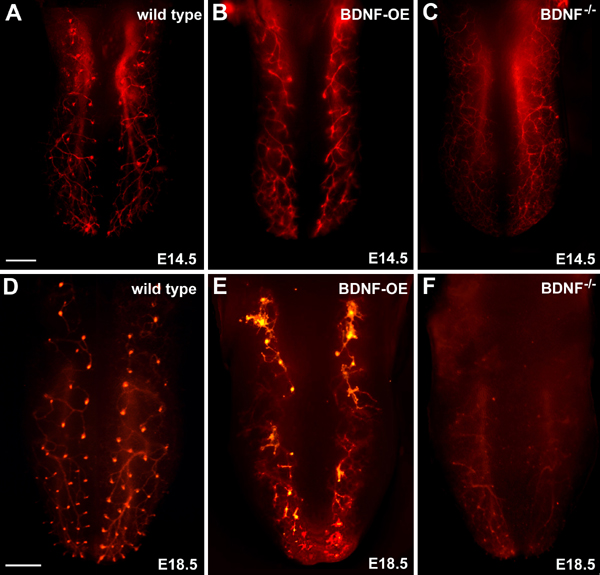
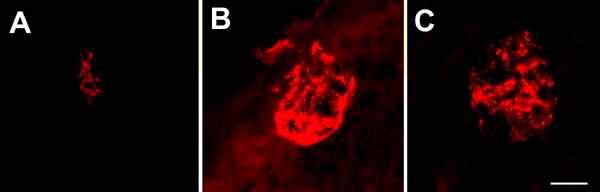
Numerous molecular factors orchestrate the development of the peripheral taste system. The unique anatomy/function of the taste system makes this system ideal for understanding the mechanisms by which these factors function; yet the taste system is underutilized for this role. This review focuses on some of the many factors that are known to regulate gustatory development, and discusses a few topics where more work is needed. Some attention is given to factors that regulate epibranchial placode formation, since gustatory neurons are thought to be primarily derived from this region. Epibranchial placodes appear to arise from a pan-placodal region and a number of regulatory factors control the differentiation of individual placodes. Gustatory neuron differentiation is regulated by a series of transcription factors and perhaps bone morphongenic proteins (BMP). As neurons differentiate, they also proliferate such that their numbers exceed those in the adult, and this is followed by developmental death. Some of these cell-cycling events are regulated by neurotrophins. After gustatory neurons become post-mitotic, axon outgrowth occurs. Axons are guided by multiple chemoattractive and chemorepulsive factors, including semaphorins, to the tongue epithelium. Brain derived neurotrophic factor (BDNF), functions as a targeting factor in the final stages of axon guidance and is required for gustatory axons to find and innervate taste epithelium. Numerous factors are involved in the development of gustatory papillae including Sox-2, Sonic hedge hog and Wnt-beta-catenin signaling. It is likely that just as many factors regulate taste bud differentiation; however, these factors have not yet been identified. Studies examining the molecular factors that regulate terminal field formation in the nucleus of the solitary tract are also lacking. However, it is possible that some of the factors that regulate geniculate ganglion development, outgrowth, guidance and targeting of peripheral axons may have the same functions in the gustatory CNS.
Figures
References
-
- Barlow LA. Gustatory System Development. In: Finger TE, Silver WL, Restrepo D, editor. The Neurobiology of Taste and Smell. New York: Wiley-Liss; 2000. pp. 395–422.
-
- Mistretta CM, Hill DL. Development of the taste system: basic neurobiology. In: RL D, editor. Handbook of Olfaction and Gustation. New York: Marcel Dekker; 1995. pp. 635–668.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
