

Review
doi: 10.1016/j.yfrne.2007.07.003.
Epub 2007 Aug 24.
Sex differences in drug abuse
Affiliations
- PMID: 17904621
- PMCID: PMC2235192
- DOI: 10.1016/j.yfrne.2007.07.003
Item in Clipboard
Review
Sex differences in drug abuse
Front Neuroendocrinol.
2008 Jan.
Abstract
Sex differences are present for all of the phases of drug abuse (initiation, escalation of use, addiction, and relapse following abstinence). While there are some differences among specific classes of abused drugs, the general pattern of sex differences is the same for all drugs of abuse. Females begin regularly self-administering licit and illicit drugs of abuse at lower doses than do males, use escalates more rapidly to addiction, and females are at greater risk for relapse following abstinence. In this review, sex differences in drug abuse are discussed for humans and in animal models. The possible neuroendocrine mechanisms mediating these sex differences are discussed.
Figures

Ovariectomized (OVX) female rats were prepared for self-administration as previously described [62]. Rats were trained on an FR1 and then an FR2 schedule of reinforcement while receiving 0.4 mg/kg/infusion cocaine, and then were transferred to a progressive ratio (PR) schedule as described by Roberts [104]. Animals were assigned to one of 2 groups: 1) OVX treated with oil 30 min prior to the PR test session for 5 days then no treatment for 2 days (OVX+OIL, N=9); or 2) OVX rats treated with 5 μg EB 30 min prior to the PR session for the first 5 days, and then no treatment for 2 days (OVX+EB, N=12). The Mean +/− SEM number of responses per 4 h progressive ratio test session is illustrated. During the first week the cocaine dose received was 0.3 mg/kg/infusion, during the second week 0.4 mg/kg/infusion, and during the last week 0.5 mg/kg/infusion. Analysis of variance with repeated measures was conducted at each dose. At 0.3 mg/kg/infusion, there was no difference between the groups. At 0.4 mg/kg/infusion, there was a main effect of Group, F1, 19=12.27, p<0.0024; an effect of Day, F4, 19=2.92, p<0.0026; and a Group by Day interaction, F4, 19=3.53, p<0.011. Post hoc pair wise comparisons indicated that OVX+ EB > OVX+OIL group (p<0.001). At 0.5 mg/kg/infusion of cocaine, there was a main effect of Group, F1 , 19=4.84, p<0.04; an effect of Day, F4 ,19=6.08, p<0.0003; but no Group by Day interaction, F4 ,19=0.25. Post hoc pair wise comparisons indicated that OVX+ EB> OVX+OIL group (p<0.001). There were no group differences in the number of nose pokes in the inactive hole during any test sessions. *denotes a significant effect of EB on progressive ratio responding as indicated above.
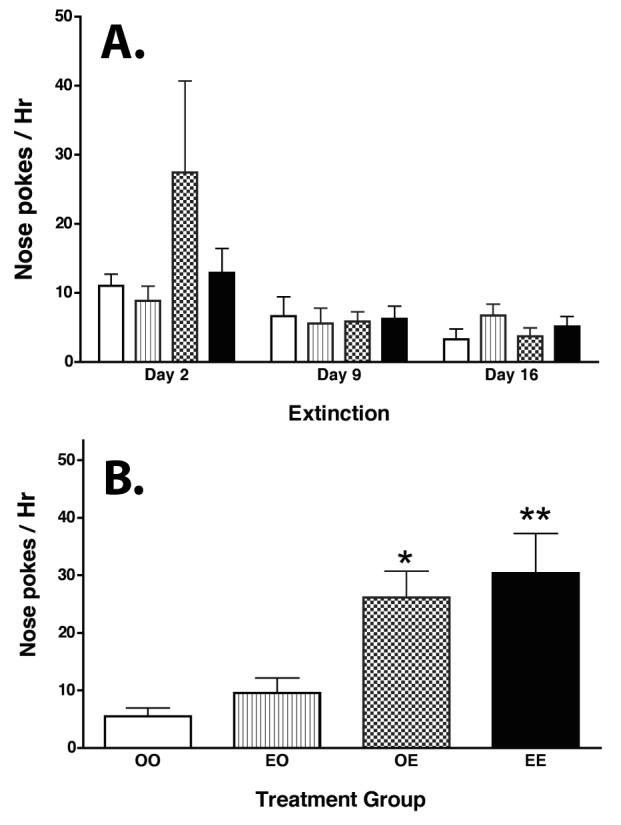
OVX rats were trained to self-administer cocaine with 0.4 mg/kg/infusion cocaine under an FR1 schedule beginning 5 days after catheter surgery using procedures described previously [62]. Animals received 0.1 ml of peanut oil or 5 μg estradiol benzoate (EB, s.c.) in 0.1 μl peanut oil (OIL, s.c.) 30 min before a three-hour self-administration training period. During this three hour training period, the house light and similar paired light + tone conditioned stimuli were present and the syringe was activated to deliver 50 μl of cocaine at the appropriate dose. Daily 3-hour sessions of self-administration training were given for 5 consecutive days followed by 2 days off for 2 weeks, animals were required to earn at least 50 infusions per day for the last two consecutive sessions to continue in the experiment. The extinction training was under an FR1 schedule on days 2, 9, 16 and 30 post-training. During this four hour extinction period, the same house light and similar paired light + tone conditioned stimuli were present and the syringe was activated to deliver 50 μl saline. There were no significant differences in the number of infusions received by the different groups during self-administration training and there were no differences among the groups during extinction. A. On days 2, 9, and 16 after the extinction session, animals received OIL or EB (half of each group received each treatment for a total of 4 groups: OIL during training + OIL during extinction testing [OIL+OIL]; OIL during training + EB during extinction testing [OIL+EB]; EB during training + OIL during extinction testing [EB+OIL]; EB during training + EB during extinction testing [EB+EB]) and were returned to the test chamber for 90 min. No cues were present during the first 30 min, then nose pokes again activated the light+tone and syringe and nose pokes were recorded for 1 hour. Open bars = OVX Oil+Oil, N=11; Gray bars = OVX Oil+E, N=7; Stippled bars = OVX EB+Oil, N=7; and Black bars = OVX EB+EB N=8. There were no significant differences in the number of active responses made by animals in these groups after EB priming. B. On day 30 of extinction, animals received OIL or EB (same groups as above) and were returned to the test chamber. Thirty minutes later animals received 5 mg/kg cocaine and nose pokes were recorded for 1 hour. Open bars = OO: OVXOil+Oil, N=11; Gray bars = EO: OVXOil+EB, N=7; Stippled bars = OE: OVXEB+Oil, N=7; and Black bars = EE: OVXEB+EB N=8. There was a significant difference in the number of active responses after treatment with 5 mg/kg cocaine (F 3, 36 =4.523, P<0.0086). Subsequent pair wise comparisons indicated that EB treatment prior to cocaine increased responding for cocaine (OVXOil+EB > OVXOil+Oil (P< 0.0294); OVXEB+EB > OVXEB+Oil (P< 0.0018); and OVXEB+EB > OVXOil+Oil (P< 0.0126). ** OVX EB+EB > OVX Oil+Oil and OVX EB+Oil (P <0.01). * OVX Oil+EB > OVX Oil+Oil rats (P <0.03)..
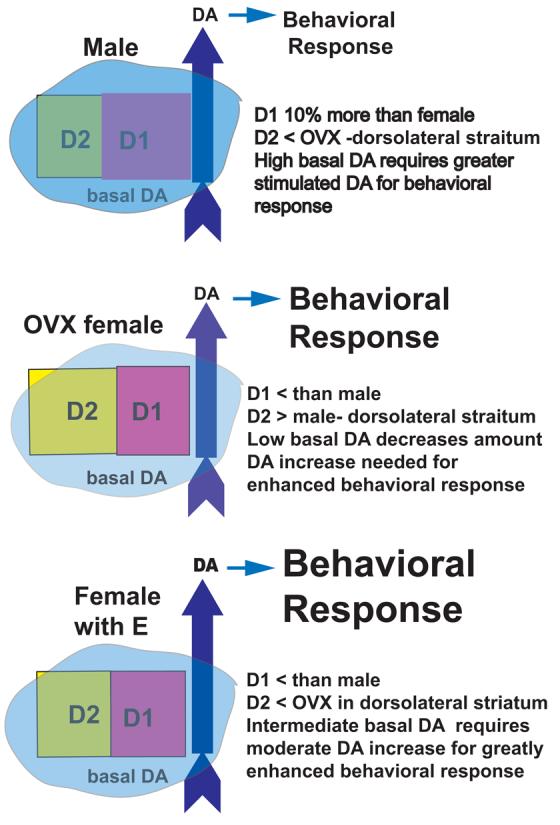
The pink and yellow squares represent the quantity of D1 and D2 DA receptors (respectively). There are 10% more D1 DA receptors (Pink Squares) in the striatum and NAcc of male rats than in intact female rats [2,57]. We find greater D2 binding in the dorsolateral striatum of OVX compared to CAST [8], while others have reported no sex differences in D2 binding when the entire striatum is considered [2,57]. Additionally, in females, estradiol rapidly down-regulates D2 DA receptor binding in dorsolateral striatum of female rats [8]. Research from the pair-bonding literature and the drug abuse literature, suggests that D1 receptors decrease affiliative behavior and addictive behavior, while D2 receptors increase these behaviors [4,112]. These D1/D2 effects are thought to be mediated by DA receptors in the NAcc. Here it is proposed that there is a similar relationship between D1/D2 receptors in dorsolateral striatum, that perhaps in concert with the NAcc, contributes to the behavioral effects and addiction liability of psychomotor stimulant drugs. The blue overlay represents the basal DA “tone”, the intensity of the color represents the quantity of basal DA, changing the color of DA receptors as their set-point for activation is attenuated by tonic DA stimulation. Results from in vivo microdialysis in freely moving rats have found that the basal extracellular concentrations of DA, as determined by the no net flux method, are twice as high in striatum of CAST males as in OVX females. Additionally, basal DA is greater in estrous females than in diestrous females suggesting that estradiol enhances basal DA “tone” [138]. The stimulated release of DA is represented by the blue arrows, where the darkness of the arrow color is proportional to the amount of release (male=females>OVX in dialysis; females>males). In experiments with in vivo voltammetry, cocaine or haloperidol induce a greater increase in electrical stimulation evoked greater extracellular DA in females than in males, possibly due to greater autoreceptor control of the dopamine transporter (DAT) [133,134]. In vitro, the AMPH-stimulated increase in striatal DA release is comparable for tissue from intact male rats and intact female rats in estrus, and release is attenuated after OVX [17]. The consequence of the coordinated effect of DA stimulation and receptor activation is a behavioral response/ rewarding effect that is greatest in females with OVX+E (or intact females in estrus)>OVX>males.
References
-
- Alele PE, Devaud LL. Sex differences in steroid modulation of ethanol withdrawal in male and female rats. Journal of Pharmacology & Experimental Therapeutics. 2007;320:427–36. - PubMed
-
- Andersen SL, Rutstein M, Benzo JM, Hostetter JC, Teicher MH. Sex differences in dopamine receptor overproduction and elimination. Neuroreport. 1997;8:1495–8. - PubMed
-
- Anglin MD, Hser YI, McGlothlin WH. Sex differences in addict careers. 2. Becoming addicted. American Journal of Drug & Alcohol Abuse. 1987;13:59–71. - PubMed
-
- Aragona BJ, Liu Y, Yu YJ, Curtis JT, Detwiler JM, Insel TR, Wang Z. Nucleus accumbens dopamine differentially mediates the formation and maintenance of monogamous pair bonds.[see comment] Nature Neuroscience. 2006;9:133–9. - PubMed
-
- Arnold AP, Gorski RA. Gonadal steroid induction of structural sex differences in the central nervous system. Annu Rev Neurosci. 1984;7:413–42. - PubMed
