

STIM2 protein mediates distinct store-dependent and store-independent modes of CRAC channel activation
- PMID: 17905723
- PMCID: PMC3601890
- DOI: 10.1096/fj.07-9449com
STIM2 protein mediates distinct store-dependent and store-independent modes of CRAC channel activation
Abstract
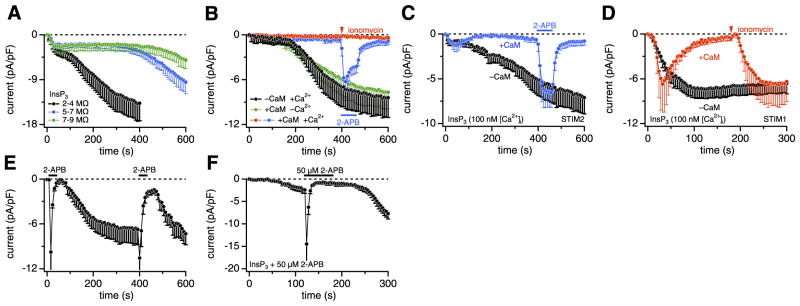
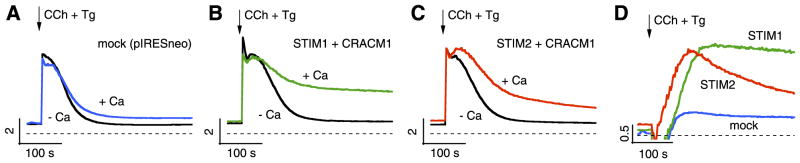
STIM1 and CRACM1 (or Orai1) are essential molecular components mediating store-operated Ca2+ entry (SOCE) and Ca2+ release-activated Ca2+ (CRAC) currents. Although STIM1 acts as a luminal Ca2+ sensor in the endoplasmic reticulum (ER), the function of STIM2 remains unclear. Here we reveal that STIM2 has two distinct modes of activating CRAC channels: a store-operated mode that is activated through depletion of ER Ca2+ stores by inositol 1,4,5-trisphosphate (InsP3) and store-independent activation that is mediated by cell dialysis during whole-cell perfusion. Both modes are regulated by calmodulin (CaM). The store-operated mode is transient in intact cells, possibly reflecting recruitment of CaM, whereas loss of CaM in perfused cells accounts for the persistence of the store-independent mode. The inhibition by CaM can be reversed by 2-aminoethoxydiphenyl borate (2-APB), resulting in rapid, store-independent activation of CRAC channels. The aminoglycoside antibiotic G418 is a highly specific and potent inhibitor of STIM2-dependent CRAC channel activation. The results reveal a novel bimodal control of CRAC channels by STIM2, the store dependence and CaM regulation, which indicates that the STIM2/CRACM1 complex may be under the control of both luminal and cytoplasmic Ca2+ levels.
Figures
Similar articles
-
STIM1 and STIM2 proteins differently regulate endogenous store-operated channels in HEK293 cells.J Biol Chem. 2015 Feb 20;290(8):4717-4727. doi: 10.1074/jbc.M114.601856. Epub 2014 Dec 22. J Biol Chem. 2015. PMID: 25533457 Free PMC article.
-
2-Aminoethoxydiphenyl borate directly facilitates and indirectly inhibits STIM1-dependent gating of CRAC channels.J Physiol. 2008 Jul 1;586(13):3061-73. doi: 10.1113/jphysiol.2008.151365. Epub 2008 Apr 10. J Physiol. 2008. PMID: 18403424 Free PMC article.
-
STIM2 drives Ca2+ oscillations through store-operated Ca2+ entry caused by mild store depletion.J Physiol. 2013 Mar 15;591(6):1433-45. doi: 10.1113/jphysiol.2012.245399. Epub 2013 Jan 28. J Physiol. 2013. PMID: 23359669 Free PMC article.
-
CRAC channelopathies.Pflugers Arch. 2010 Jul;460(2):417-35. doi: 10.1007/s00424-009-0777-5. Epub 2010 Jan 29. Pflugers Arch. 2010. PMID: 20111871 Free PMC article. Review.
-
The neglected CRAC proteins: Orai2, Orai3, and STIM2.Curr Top Membr. 2013;71:237-71. doi: 10.1016/B978-0-12-407870-3.00010-X. Curr Top Membr. 2013. PMID: 23890118 Review.
Cited by
-
Identification of molecular determinants that govern distinct STIM2 activation dynamics.PLoS Biol. 2018 Nov 16;16(11):e2006898. doi: 10.1371/journal.pbio.2006898. eCollection 2018 Nov. PLoS Biol. 2018. PMID: 30444880 Free PMC article.
-
Stim and Orai mediate constitutive Ca2+ entry and control endoplasmic reticulum Ca2+ refilling in primary cultures of colorectal carcinoma cells.Oncotarget. 2018 Jul 24;9(57):31098-31119. doi: 10.18632/oncotarget.25785. eCollection 2018 Jul 24. Oncotarget. 2018. PMID: 30123430 Free PMC article.
-
Store-operated Ca²+ signaling in dendritic cells occurs independently of STIM1.J Leukoc Biol. 2011 Jan;89(1):57-62. doi: 10.1189/jlb.0610381. Epub 2010 Oct 22. J Leukoc Biol. 2011. PMID: 20971921 Free PMC article.
-
A cytosolic STIM2 preprotein created by signal peptide inefficiency activates ORAI1 in a store-independent manner.J Biol Chem. 2011 May 6;286(18):16174-85. doi: 10.1074/jbc.M110.206946. Epub 2011 Mar 7. J Biol Chem. 2011. PMID: 21383014 Free PMC article.
-
STIM2 (Stromal Interaction Molecule 2)-Mediated Increase in Resting Cytosolic Free Ca2+ Concentration Stimulates PASMC Proliferation in Pulmonary Arterial Hypertension.Hypertension. 2018 Mar;71(3):518-529. doi: 10.1161/HYPERTENSIONAHA.117.10503. Epub 2018 Jan 22. Hypertension. 2018. PMID: 29358461 Free PMC article.
References
-
- Parekh AB, Penner R. Store depletion and calcium influx. Physiol Rev. 1997;77:901–930. - PubMed
-
- Parekh AB, Putney JW., Jr Store-operated calcium channels. Physiol Rev. 2005;85:757– 810. - PubMed
-
- Putney JW., Jr Capacitative calcium entry revisited. Cell Calcium. 1990;11:611– 624. - PubMed
-
- Hoth M, Penner R. Depletion of intracellular calcium stores activates a calcium current in mast cells. Nature. 1992;355:353–356. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
