

Backbone-base inclination as a fundamental determinant of nucleic acid self- and cross-pairing
- PMID: 17905816
- PMCID: PMC2095819
- DOI: 10.1093/nar/gkm612
Backbone-base inclination as a fundamental determinant of nucleic acid self- and cross-pairing
Abstract
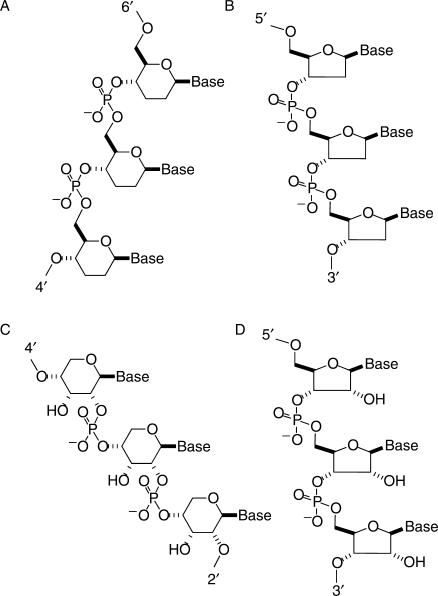
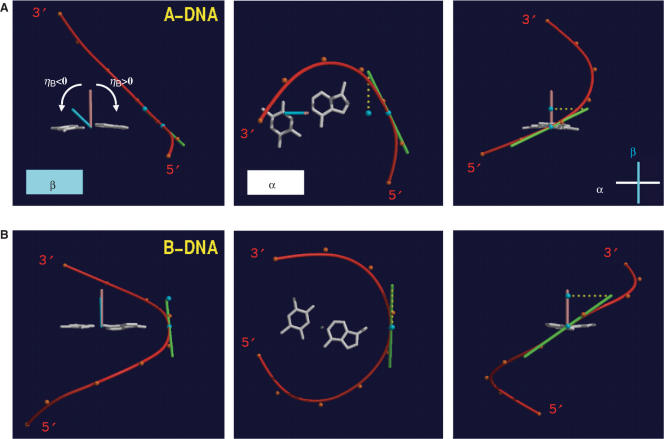
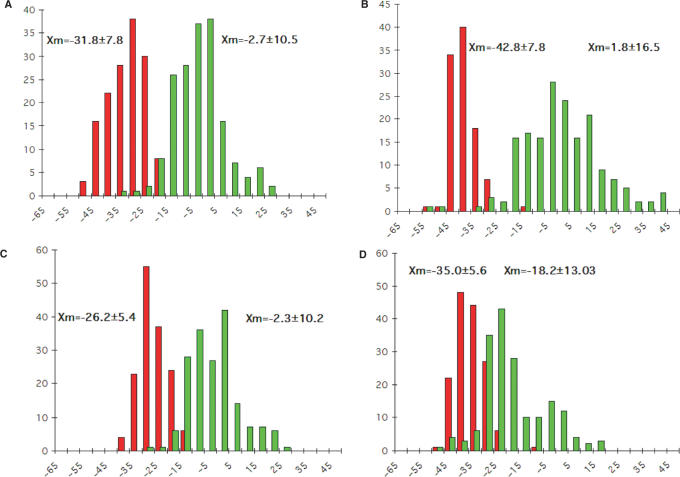
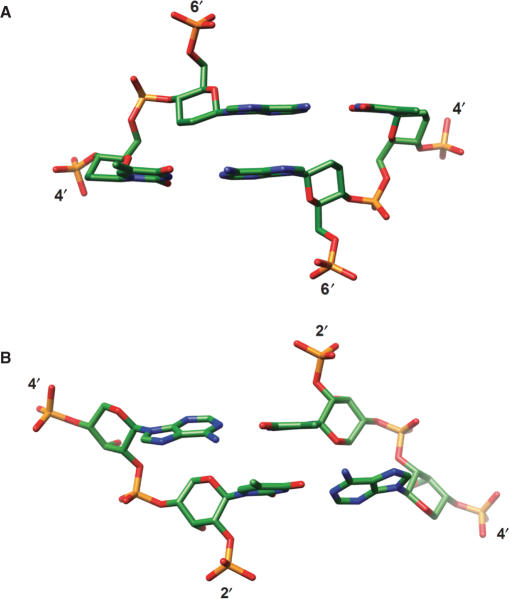
The crystal structure of the duplex formed by oligo(2',3'-dideoxy-beta-d-glucopyranosyl)nucleotides (homo-DNA) revealed strongly inclined backbone and base-pair axes [Egli,M., Pallan,P.S., Pattanayek,R., Wilds,C.J., Lubini,P., Minasov,G., Dobler,M., Leumann,C.J. and Eschenmoser,A. (2006) Crystal structure of homo-DNA and nature's choice of pentose over hexose in the genetic system. J. Am. Chem. Soc., 128, 10847-10856]. This inclination is easily perceived because homo-DNA exhibits only a modest helical twist. Conversely, the tight coiling of strands conceals that the backbone-base inclinations for A- (DNA and RNA) and B-form (DNA) duplexes differ considerably. We have defined a parameter eta(B) that corresponds to the local inclination between sugar-phosphate backbone and base plane in nucleic acid strands. Here, we show its biological significance as a predictive measure for the relative strand polarities (antiparallel, aps, or parallel, ps) in duplexes of DNA, RNA and artificial nucleic acid pairing systems. The potential of formation of ps duplexes between complementary 16-mers with eight A and U(T) residues each was investigated with DNA, RNA, 2'-O-methylated RNA, homo-DNA and p-RNA, the ribopyranosyl isomer of RNA. The thermodynamic stabilities of the corresponding aps duplexes were also measured. As shown previously, DNA is capable of forming both ps and aps duplexes. However, all other tested systems are unable to form stable ps duplexes with reverse Watson-Crick (rWC) base pairs. This observation illustrates the handicap encountered by nucleic acid systems with inclinations eta(B) that differ significantly from 0 degrees to form a ps rWC paired duplex. Accordingly, RNA with a backbone-base inclination of -30 degrees , pairs strictly in an aps fashion. On the other hand, the more or less perpendicular orientation of backbone and bases in DNA allows it to adopt a ps rWC paired duplex. In addition to providing a rationalization of relative strand polarity with nucleic acids, the backbone-base inclination parameter is also a determinant of cross-pairing. Thus, systems with strongly deviating eta(B) angles will not pair with each other. Nucleic acid pairing systems with significant backbone-base inclinations can also be expected to display different stabilities depending on which terminus carries unpaired nucleotides. The negative inclination of RNA is consistent with the higher stability of duplexes with 3'- compared to those with 5'-dangling ends.
Figures
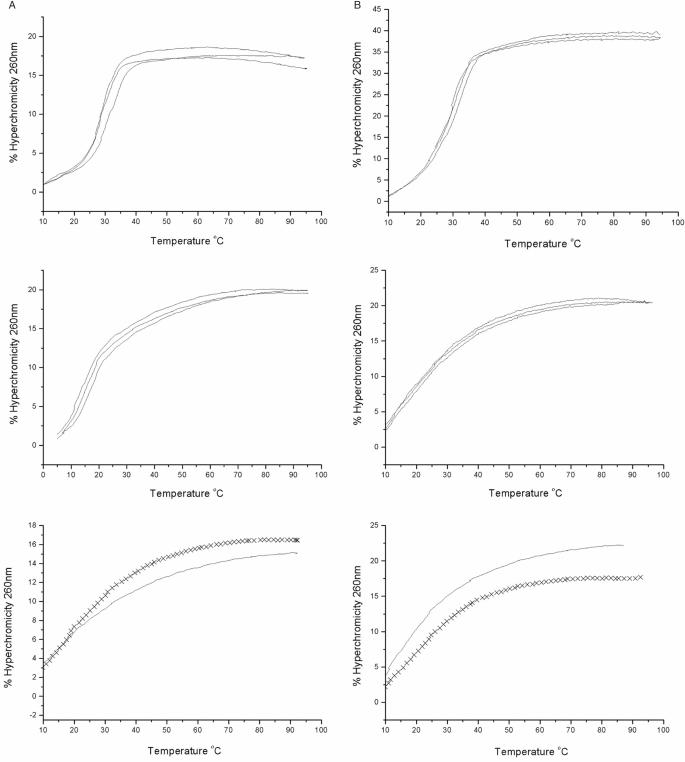
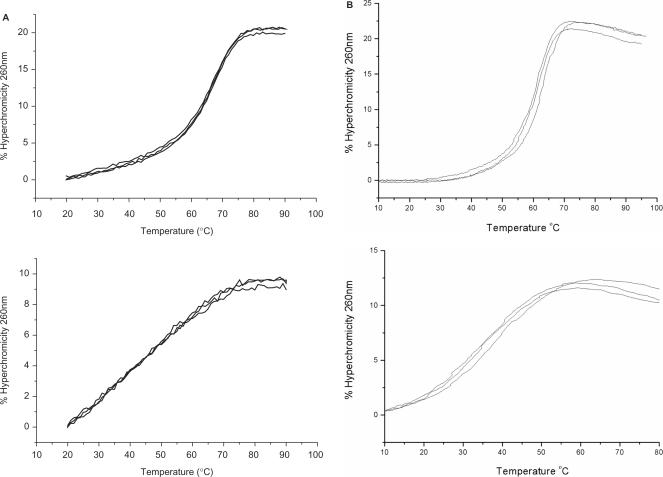
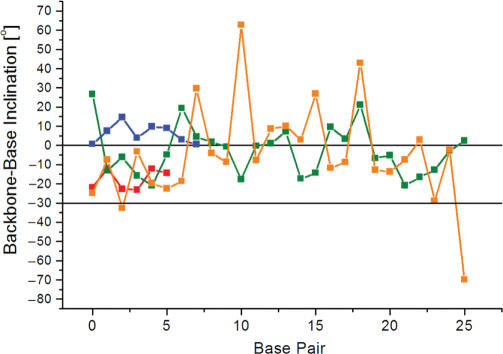
References
-
- Watson JD, Crick FHC. A structure for deoxyribose nucleic acid. Nature. 1953;171:737–738. - PubMed
-
- Franklin RE, Gosling RG. Molecular configuration in sodium thymonucleate. Nature. 1953;171:740–741. - PubMed
-
- Pattabiraman N. Can the double helix be parallel? Biopolymers. 1986;25:1603–1606. - PubMed
-
- Van de Sande JH, Ramsing NB, Germann MW, Elhorst W, Kalisch BW, von Kitzing E, Pon RT, Clegg RC, Jovin TM. Parallel stranded DNA. Science. 1988;241:551–557. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
