

Pten controls lung morphogenesis, bronchioalveolar stem cells, and onset of lung adenocarcinomas in mice
- PMID: 17909629
- PMCID: PMC1994617
- DOI: 10.1172/JCI31854
Pten controls lung morphogenesis, bronchioalveolar stem cells, and onset of lung adenocarcinomas in mice
Abstract
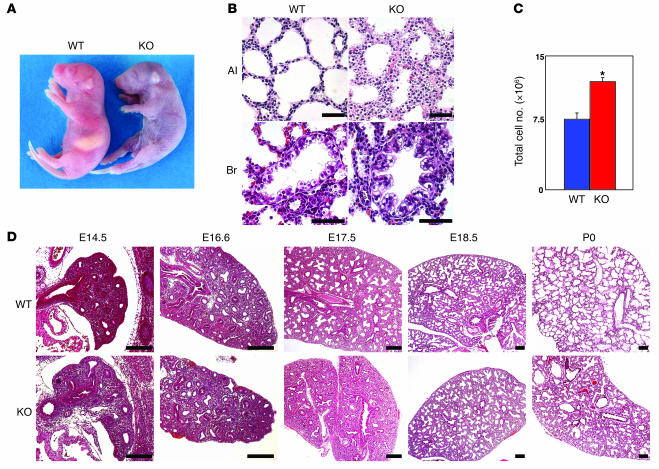
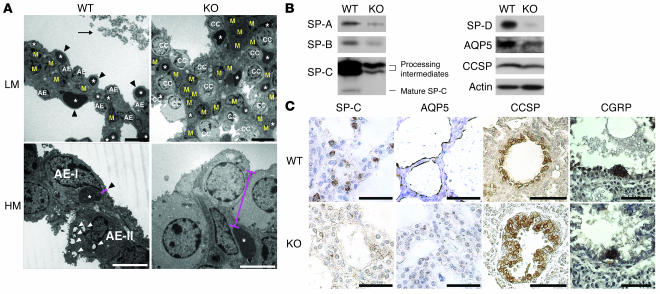
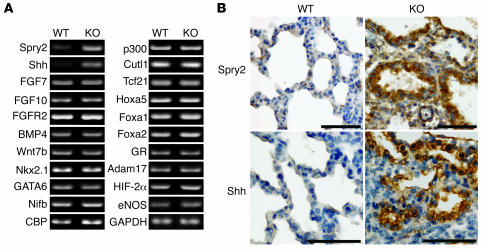
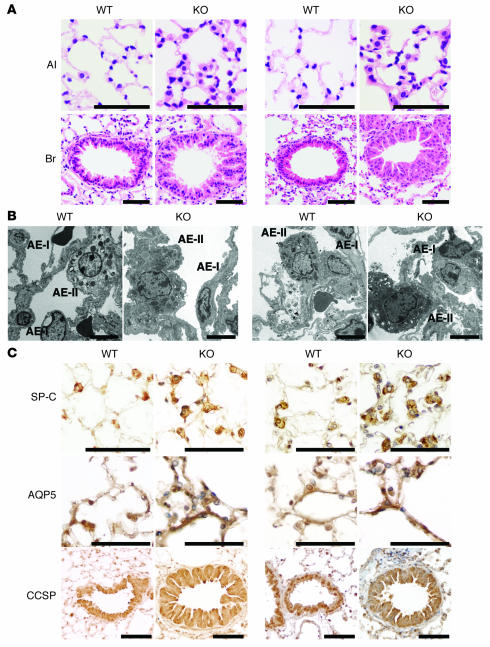
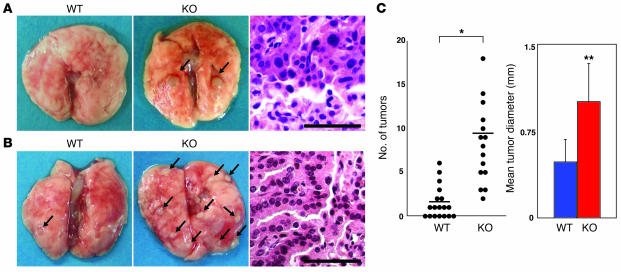
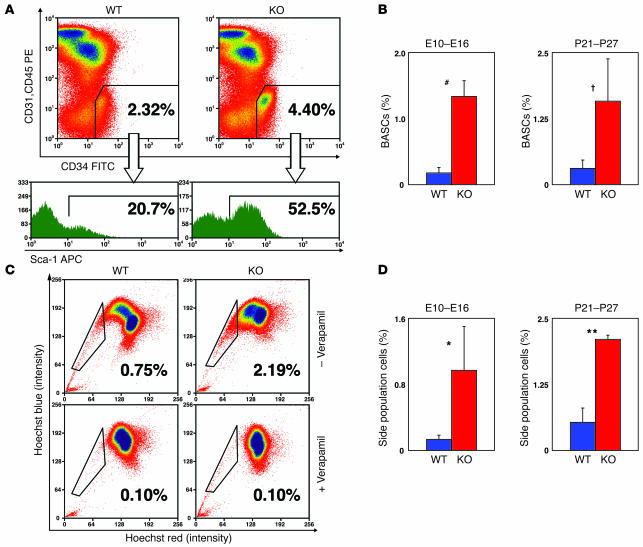
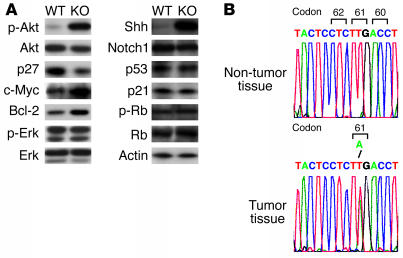
PTEN is a tumor suppressor gene mutated in many human cancers. We generated a bronchioalveolar epithelium-specific null mutation of Pten in mice [SP-C-rtTA/(tetO)(7)-Cre/Pten(flox/flox) (SOPten(flox/flox)) mice] that was under the control of doxycycline. Ninety percent of SOPten(flox/flox) mice that received doxycycline in utero [SOPten(flox/flox)(E10-16) mice] died of hypoxia soon after birth. Surviving SOPten(flox/flox)(E10-16) mice and mice that received doxycycline postnatally [SOPten(flox/flox)(P21-27) mice] developed spontaneous lung adenocarcinomas. Urethane treatment accelerated number and size of lung tumors developing in SOPten(flox/flox) mice of both ages. Histological and biochemical examinations of the lungs of SOPten(flox/flox)(E10-16) mice revealed hyperplasia of bronchioalveolar epithelial cells and myofibroblast precursors, enlarged alveolar epithelial cells, and impaired production of surfactant proteins. Numbers of bronchioalveolar stem cells (BASCs), putative initiators of lung adenocarcinomas, were increased. Lungs of SOPten(flox/flox)(E10-16) mice showed increased expression of Spry2, which inhibits the maturation of alveolar epithelial cells. Levels of Akt, c-Myc, Bcl-2, and Shh were also elevated in SOPten(flox/flox)(E10-16) and SOPten(flox/flox)(P21-27) lungs. Furthermore, K-ras was frequently mutated in adenocarcinomas observed in SOPten(flox/flox)(P21-27) lungs. These results indicate that Pten is essential for both normal lung morphogenesis and the prevention of lung carcinogenesis, possibly because this tumor suppressor is required for BASC homeostasis.
Figures
Similar articles
-
Epithelial Pten controls acute lung injury and fibrosis by regulating alveolar epithelial cell integrity.Am J Respir Crit Care Med. 2013 Feb 1;187(3):262-75. doi: 10.1164/rccm.201205-0851OC. Epub 2012 Dec 13. Am J Respir Crit Care Med. 2013. PMID: 23239155
-
MOB1-YAP1/TAZ-NKX2.1 axis controls bronchioalveolar cell differentiation, adhesion and tumour formation.Oncogene. 2017 Jul 20;36(29):4201-4211. doi: 10.1038/onc.2017.58. Epub 2017 Mar 27. Oncogene. 2017. PMID: 28346423
-
Matrix metalloproteinase-10 promotes Kras-mediated bronchio-alveolar stem cell expansion and lung cancer formation.PLoS One. 2011;6(10):e26439. doi: 10.1371/journal.pone.0026439. Epub 2011 Oct 17. PLoS One. 2011. PMID: 22022614 Free PMC article.
-
K-ras p21 expression and activity in lung and lung tumors.Exp Lung Res. 2000 Dec;26(8):659-71. doi: 10.1080/01902140150216747. Exp Lung Res. 2000. PMID: 11195463 Review.
-
Molecular pathogenesis of transplacentally induced mouse lung tumors.Exp Lung Res. 1998 Jul-Aug;24(4):557-77. doi: 10.3109/01902149809087386. Exp Lung Res. 1998. PMID: 9659583 Review.
Cited by
-
Choriodecidual infection downregulates angiogenesis and morphogenesis pathways in fetal lungs from Macaca nemestrina.PLoS One. 2012;7(10):e46863. doi: 10.1371/journal.pone.0046863. Epub 2012 Oct 9. PLoS One. 2012. PMID: 23056493 Free PMC article.
-
Loss of PTEN induces lung fibrosis via alveolar epithelial cell senescence depending on NF-κB activation.Aging Cell. 2019 Feb;18(1):e12858. doi: 10.1111/acel.12858. Epub 2018 Dec 12. Aging Cell. 2019. PMID: 30548445 Free PMC article.
-
MicroRNA-4286 promotes cell proliferation, migration, and invasion via PTEN regulation of the PI3K/Akt pathway in non-small cell lung cancer.Cancer Med. 2019 Jul;8(7):3520-3531. doi: 10.1002/cam4.2220. Epub 2019 May 10. Cancer Med. 2019. PMID: 31074594 Free PMC article.
-
Cancer stem cells: progress and challenges in lung cancer.Stem Cell Investig. 2014 Apr 15;1:9. doi: 10.3978/j.issn.2306-9759.2014.03.06. eCollection 2014. Stem Cell Investig. 2014. PMID: 27358855 Free PMC article. Review.
-
Isolation of alveolar epithelial type II progenitor cells from adult human lungs.Lab Invest. 2011 Mar;91(3):363-78. doi: 10.1038/labinvest.2010.187. Epub 2010 Nov 15. Lab Invest. 2011. PMID: 21079581 Free PMC article. Review.
References
-
- Jemal A., Thomas A., Murray T., Thun M. Cancer statistics, 2002. CA Cancer J. Clin. 2002;52:23–47. - PubMed
-
- Giangreco A., Groot K.R., Janes S.M. Lung cancer and lung stem cells: strange bedfellows? Am. J. Respir. Crit. Care Med. 2007;175:547–553. - PubMed
-
- Minna J.D., Roth J.A., Gazdar A.F. Focus on lung cancer. Cancer Cell. 2002;1:49–52. - PubMed
-
- Reya T., Morrison S.J., Clarke M.F., Weissman I.L. Stem cells, cancer, and cancer stem cells. Nature. 2001;414:105–111. - PubMed
-
- Kim C.F., et al. Identification of bronchioalveolar stem cells in normal lung and lung cancer. Cell. 2005;121:823–835. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
