

Silent development of memory progenitor B cells
- PMID: 17911603
- PMCID: PMC2896480
- DOI: 10.4049/jimmunol.179.8.5181
Silent development of memory progenitor B cells
Abstract
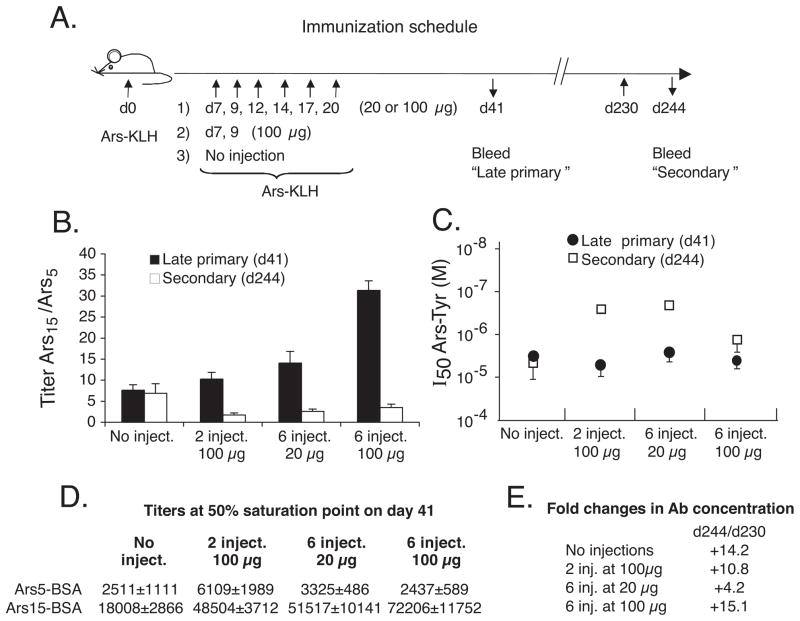
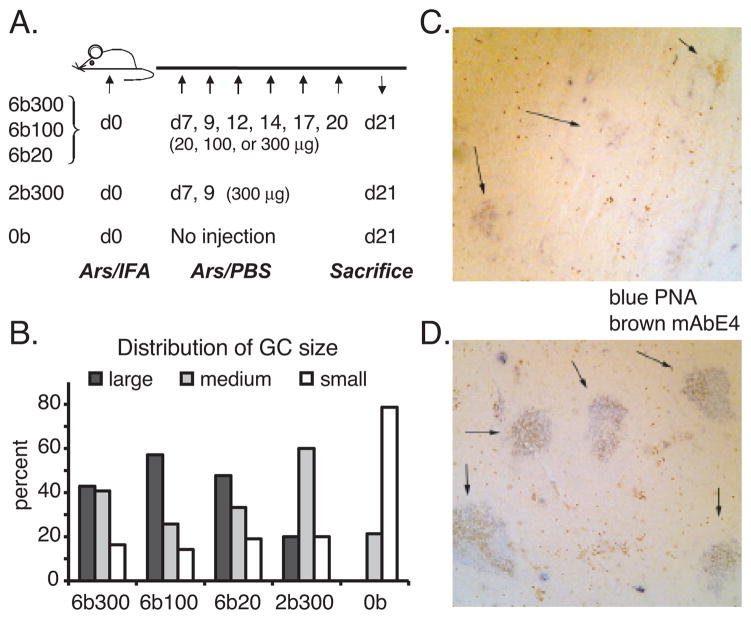
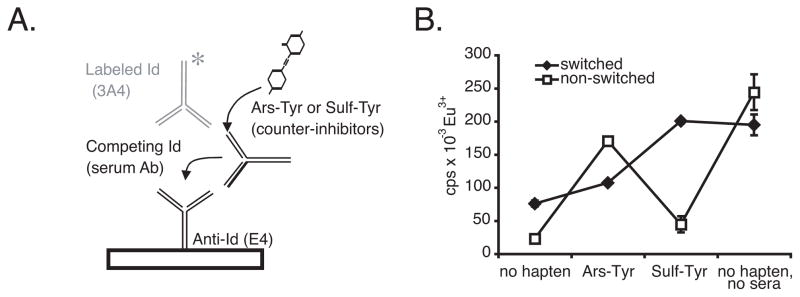
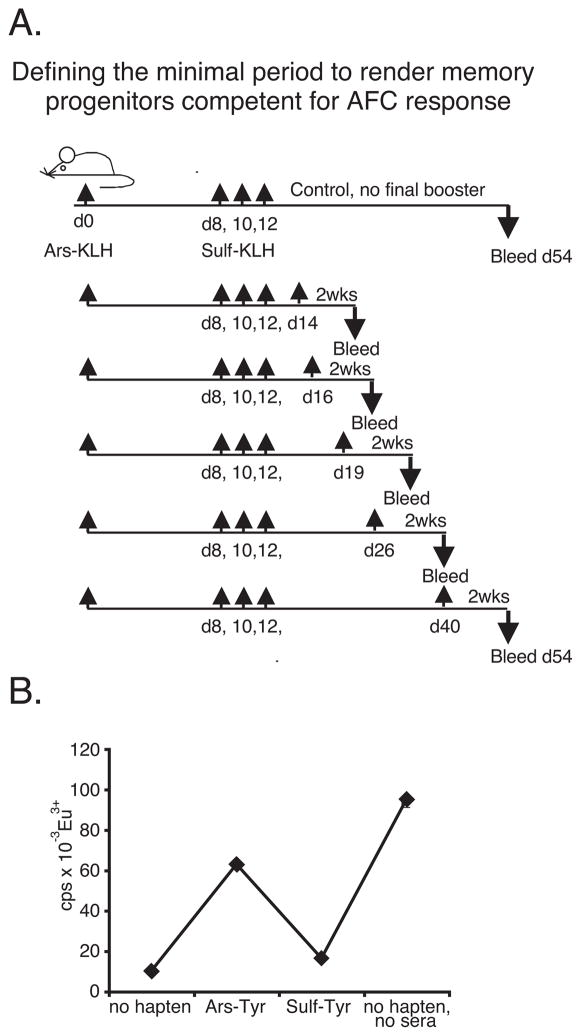
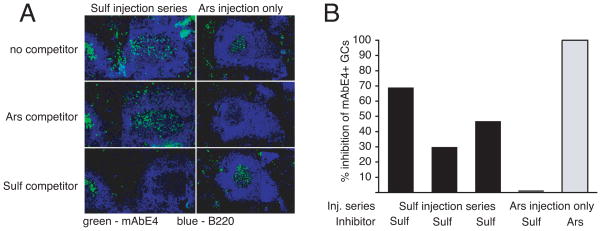
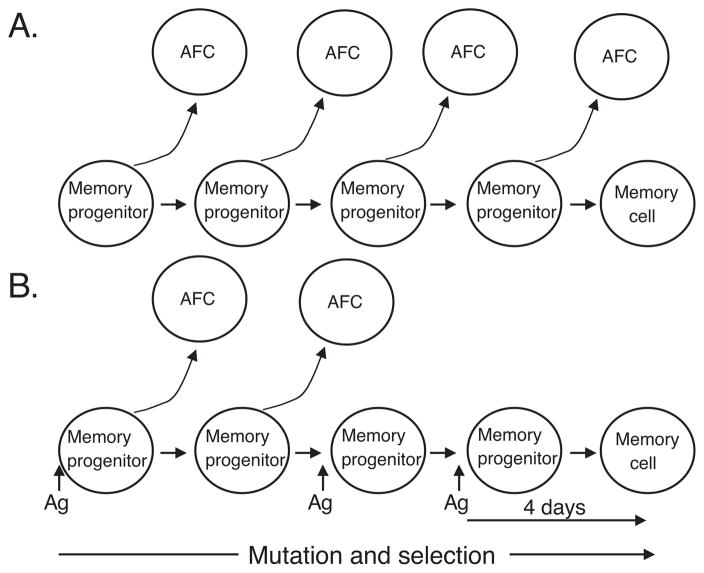
T cell-dependent immune responses generate long-lived plasma cells and memory B cells, both of which express hypermutated Ab genes. The relationship between these cell types is not entirely understood. Both appear to emanate from the germinal center reaction, but it is unclear whether memory cells evolve while obligatorily generating plasma cells by siblings under all circumstances. In the experiments we report, plasma cell development was functionally segregated from memory cell development by a series of closely spaced injections of Ag delivered during the period of germinal center development. The injection series elevated serum Ab of low affinity, supporting the idea that a strong Ag signal drives plasma cell development. At the same time, the injection series produced a distinct population of affinity/specificity matured memory B cells that were functionally silent, as manifested by an absence of corresponding serum Ab. These cells could be driven by a final booster injection to develop into Ab-forming cells. This recall response required that a rest period precede the final booster injection, but a pause of only 4 days was sufficient. Our results support a model of memory B cell development in which extensive affinity/specificity maturation can take place within a B cell clone under some circumstances in which a concomitant generation of Ab-forming cells by siblings does not take place.
Figures
Similar articles
-
Commitment of B lymphocytes to a plasma cell fate is associated with Blimp-1 expression in vivo.J Immunol. 2000 Nov 15;165(10):5462-71. doi: 10.4049/jimmunol.165.10.5462. J Immunol. 2000. PMID: 11067898
-
Impaired affinity maturation in Cr2-/- mice is rescued by adjuvants without improvement in germinal center development.J Immunol. 2000 Sep 15;165(6):3119-27. doi: 10.4049/jimmunol.165.6.3119. J Immunol. 2000. PMID: 10975825
-
IL-21 receptor is critical for the development of memory B cell responses.J Immunol. 2011 Jan 15;186(2):667-74. doi: 10.4049/jimmunol.0903207. Epub 2010 Dec 17. J Immunol. 2011. PMID: 21169545
-
Plasticity and heterogeneity in the generation of memory B cells and long-lived plasma cells: the influence of germinal center interactions and dynamics.J Immunol. 2010 Sep 15;185(6):3117-25. doi: 10.4049/jimmunol.1001155. J Immunol. 2010. PMID: 20814029 Review.
-
The generation of memory B cells.Semin Immunol. 1992 Feb;4(1):3-9. Semin Immunol. 1992. PMID: 1591368 Review.
Cited by
-
Aborted germinal center reactions and B cell memory by follicular T cells specific for a B cell receptor V region peptide.J Immunol. 2011 Jul 1;187(1):212-21. doi: 10.4049/jimmunol.1002328. Epub 2011 May 27. J Immunol. 2011. PMID: 21622866 Free PMC article.
-
Cancer stem cells in multiple myeloma.Cancer Lett. 2009 May 8;277(1):1-7. doi: 10.1016/j.canlet.2008.08.005. Epub 2008 Sep 21. Cancer Lett. 2009. PMID: 18809245 Free PMC article. Review.
References
-
- MacLennan IC, Gray D. Antigen-driven selection of virgin and memory B cells. Immunol Rev. 1986;91:61–85. - PubMed
-
- Vieira P, Rajewsky K. Persistence of memory B cells in mice deprived of T cell help. Int Immunol. 1990;2:487–494. - PubMed
-
- Manz RA, Thiel A, Radbruch A. Lifetime of plasma cells in the bone marrow. Nature. 1997;388:133–134. - PubMed
-
- Slifka MK, Ahmed R. Long-lived plasma cells: a mechanism for maintaining persistent antibody production. Curr Opin Immunol. 1998;10:252–258. - PubMed
-
- Tarlinton D. Germinal centers: form and function. Curr Opin Immunol. 1998;10:245–251. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
