

Structure and reaction mechanism of basil eugenol synthase
- PMID: 17912370
- PMCID: PMC1991597
- DOI: 10.1371/journal.pone.0000993
Structure and reaction mechanism of basil eugenol synthase
Abstract
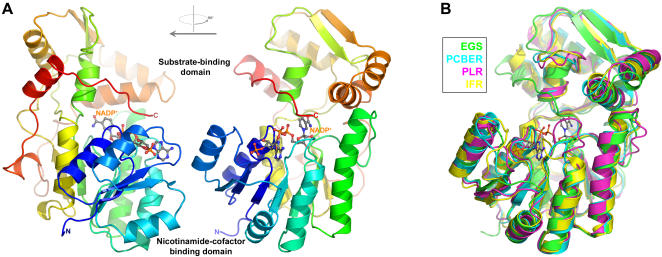
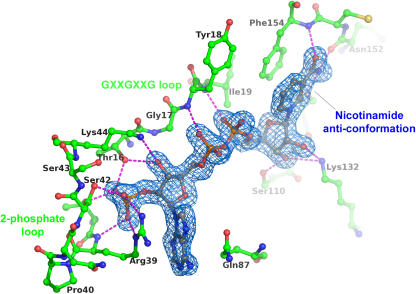
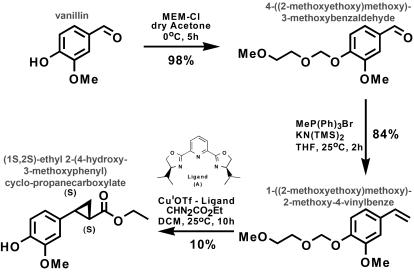
Phenylpropenes, a large group of plant volatile compounds that serve in multiple roles in defense and pollinator attraction, contain a propenyl side chain. Eugenol synthase (EGS) catalyzes the reductive displacement of acetate from the propenyl side chain of the substrate coniferyl acetate to produce the allyl-phenylpropene eugenol. We report here the structure determination of EGS from basil (Ocimum basilicum) by protein x-ray crystallography. EGS is structurally related to the short-chain dehydrogenase/reductases (SDRs), and in particular, enzymes in the isoflavone-reductase-like subfamily. The structure of a ternary complex of EGS bound to the cofactor NADP(H) and a mixed competitive inhibitor EMDF ((7S,8S)-ethyl (7,8-methylene)-dihydroferulate) provides a detailed view of the binding interactions within the EGS active site and a starting point for mutagenic examination of the unusual reductive mechanism of EGS. The key interactions between EMDF and the EGS-holoenzyme include stacking of the phenyl ring of EMDF against the cofactor's nicotinamide ring and a water-mediated hydrogen-bonding interaction between the EMDF 4-hydroxy group and the side-chain amino moiety of a conserved lysine residue, Lys132. The C4 carbon of nicotinamide resides immediately adjacent to the site of hydride addition, the C7 carbon of cinnamyl acetate substrates. The inhibitor-bound EGS structure suggests a two-step reaction mechanism involving the formation of a quinone-methide prior to reduction. The formation of this intermediate is promoted by a hydrogen-bonding network that favors deprotonation of the substrate's 4-hydroxyl group and disfavors binding of the acetate moiety, akin to a push-pull catalytic mechanism. Notably, the catalytic involvement in EGS of the conserved Lys132 in preparing the phenolic substrate for quinone methide formation through the proton-relay network appears to be an adaptation of the analogous role in hydrogen bonding played by the equivalent lysine residue in other enzymes of the SDR family.
Conflict of interest statement
Figures






References
-
- Dexter R, Qualley A, Kish CM, Ma CJ, Koeduka T, et al. Characterization of a petunia acetyltransferase involved in the biosynthesis of the floral volatile isoeugenol. Plant J. 2007;49:265–275. - PubMed
-
- Vassao DG, Gang DR, Koeduka T, Jackson B, Pichersky E, et al. Chavicol formation in sweet basil (Ocimum basilicum): cleavage of an esterified C9 hydroxyl group with NAD(P)H-dependent reduction. Org Biomol Chem. 2006;4:2733–2744. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
