

Identification of regions and residues in feline junctional adhesion molecule required for feline calicivirus binding and infection
- PMID: 17913818
- PMCID: PMC2168846
- DOI: 10.1128/JVI.01509-07
Identification of regions and residues in feline junctional adhesion molecule required for feline calicivirus binding and infection
Abstract
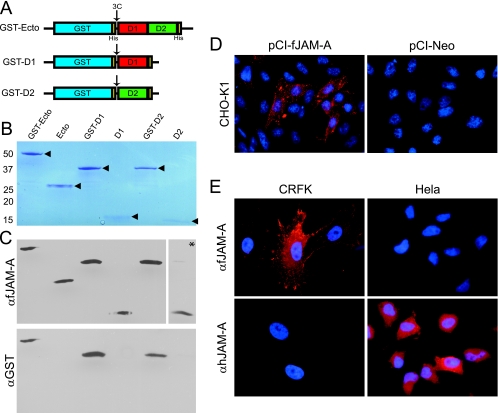
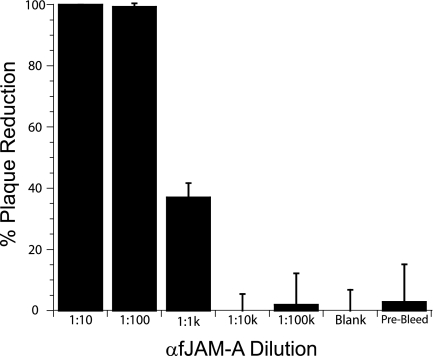
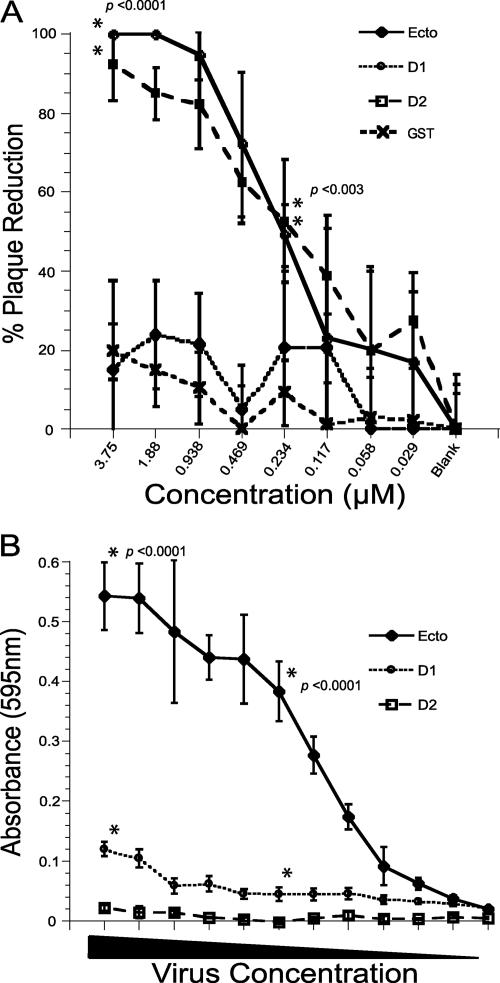
The feline junctional adhesion molecule A (fJAM-A) is a functional receptor for feline calicivirus (FCV). fJAM-A is a member of the immunoglobulin superfamily (IgSF) and consists of two Ig-like extracellular domains (D1 and D2), a membrane-spanning domain, and a short cytoplasmic tail. To identify regions of fJAM-A that interact with FCV, we purified recombinant fJAM-A ectodomain and D1 and D2 domains. We found that preincubation of FCV with the ectodomain or D1 was sufficient to inhibit FCV infection in plaque reduction assays. In enzyme-linked immunosorbent assays, FCV binding to fJAM-A ectodomain was concentration dependent and saturable; however, FCV bound D1 alone weakly and was unable to bind D2. To characterize FCV binding to surface-expressed fJAM-A, we transfected truncated and chimeric forms of fJAM-A into a nonpermissive cell line and assayed binding by flow cytometry. Only D1 was necessary for FCV binding to cells; all other domains could be replaced. Using a structure-guided mutational approach, we identified three mutants of fJAM-A within D1 (D42N, K43N, and S97A) that exhibited significantly decreased capacities to bind FCV. In contrast to our finding that D1 mediated FCV binding, we found that all domains of fJAM-A were necessary to confer susceptibility to FCV infection. Furthermore, surface expression of fJAM-A was not sufficient to permit FCV infection by all of the isolates we investigated. This indicates that (i) other cellular factors are required to permit productive FCV infection and (ii) individual FCV isolates differ in the factors they require.
Figures
Similar articles
-
Conserved Surface Residues on the Feline Calicivirus Capsid Are Essential for Interaction with Its Receptor Feline Junctional Adhesion Molecule A (fJAM-A).J Virol. 2018 Mar 28;92(8):e00035-18. doi: 10.1128/JVI.00035-18. Print 2018 Apr 15. J Virol. 2018. PMID: 29386293 Free PMC article.
-
Distribution of the feline calicivirus receptor junctional adhesion molecule a in feline tissues.Vet Pathol. 2011 Mar;48(2):361-8. doi: 10.1177/0300985810375245. Epub 2010 Jul 15. Vet Pathol. 2011. PMID: 20634409
-
Junctional adhesion molecule 1 is a functional receptor for feline calicivirus.J Virol. 2006 May;80(9):4482-90. doi: 10.1128/JVI.80.9.4482-4490.2006. J Virol. 2006. PMID: 16611908 Free PMC article.
-
[Progress in establishment and application of feline calicivirus reverse genetics operating system].Bing Du Xue Bao. 2015 Jan;31(1):74-9. Bing Du Xue Bao. 2015. PMID: 25997334 Review. Chinese.
-
Expression cloning of signaling proteins regulated by cell adhesion.Methods Mol Biol. 2006;341:155-65. doi: 10.1385/1-59745-113-4:155. Methods Mol Biol. 2006. PMID: 16799197 Review.
Cited by
-
Norovirus Attachment and Entry.Viruses. 2019 May 30;11(6):495. doi: 10.3390/v11060495. Viruses. 2019. PMID: 31151248 Free PMC article. Review.
-
Fexaramine as an entry blocker for feline caliciviruses.Antiviral Res. 2018 Apr;152:76-83. doi: 10.1016/j.antiviral.2018.02.009. Epub 2018 Feb 15. Antiviral Res. 2018. PMID: 29454892 Free PMC article.
-
The cryo-electron microscopy structure of feline calicivirus bound to junctional adhesion molecule A at 9-angstrom resolution reveals receptor-induced flexibility and two distinct conformational changes in the capsid protein VP1.J Virol. 2011 Nov;85(21):11381-90. doi: 10.1128/JVI.05621-11. Epub 2011 Aug 24. J Virol. 2011. PMID: 21865392 Free PMC article.
-
Multiple Correspondence Analysis on Amino Acid Properties within the Variable Region of the Capsid Protein Shows Differences between Classical and Virulent Systemic Feline Calicivirus Strains.Viruses. 2019 Nov 23;11(12):1090. doi: 10.3390/v11121090. Viruses. 2019. PMID: 31771183 Free PMC article.
-
Identification of Human Junctional Adhesion Molecule 1 as a Functional Receptor for the Hom-1 Calicivirus on Human Cells.mBio. 2017 Feb 14;8(1):e00031-17. doi: 10.1128/mBio.00031-17. mBio. 2017. PMID: 28196955 Free PMC article.
References
-
- Babinska, A., M. H. Kedees, H. Athar, T. Ahmed, O. Batuman, Y. H. Ehrlich, M. M. Hussain, and E. Kornecki. 2002. F11-receptor (F11R/JAM) mediates platelet adhesion to endothelial cells: role in inflammatory thrombosis. Thromb. Haemost. 88:843-850. - PubMed
-
- Babinska, A., M. H. Kedees, H. Athar, T. Sobocki, M. B. Sobocka, T. Ahmed, Y. H. Ehrlich, M. M. Hussain, and E. Kornecki. 2002. Two regions of the human platelet F11-receptor (F11R) are critical for platelet aggregation, potentiation and adhesion. Thromb. Haemost. 87:712-721. - PubMed
-
- Baldwin, S. L., T. D. Powell, K. S. Sellins, S. V. Radecki, J. John Cohen, and M. J. Milhausen. 2004. The biological effects of five feline IFN-alpha subtypes. Vet. Immunol. Immunopathol. 99:153-167. - PubMed
-
- Barton, E. S., J. C. Forrest, J. L. Connolly, J. D. Chappell, Y. Liu, F. J. Schnell, A. Nusrat, C. A. Parkos, and T. S. Dermody. 2001. Junction adhesion molecule is a receptor for reovirus. Cell 104:441-451. - PubMed
-
- Bazzoni, G., O. M. Martinez-Estrada, F. Orsenigo, M. Cordenonsi, S. Citi, and E. Dejana. 2000. Interaction of junctional adhesion molecule with the tight junction components ZO-1, cingulin, and occludin. J. Biol. Chem. 275:20520-20526. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
