

Enhanced astrocytic Ca2+ signals contribute to neuronal excitotoxicity after status epilepticus
- PMID: 17913901
- PMCID: PMC2917229
- DOI: 10.1523/JNEUROSCI.2001-07.2007
Enhanced astrocytic Ca2+ signals contribute to neuronal excitotoxicity after status epilepticus
Abstract
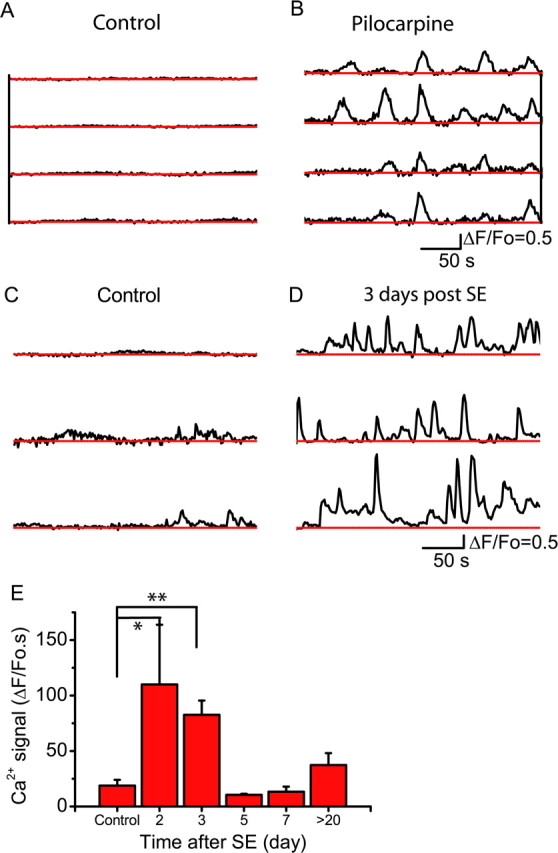
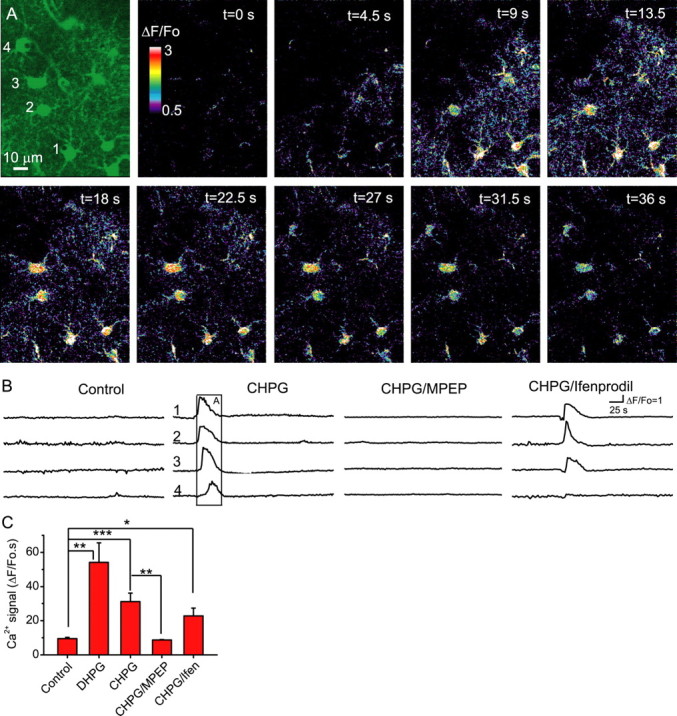
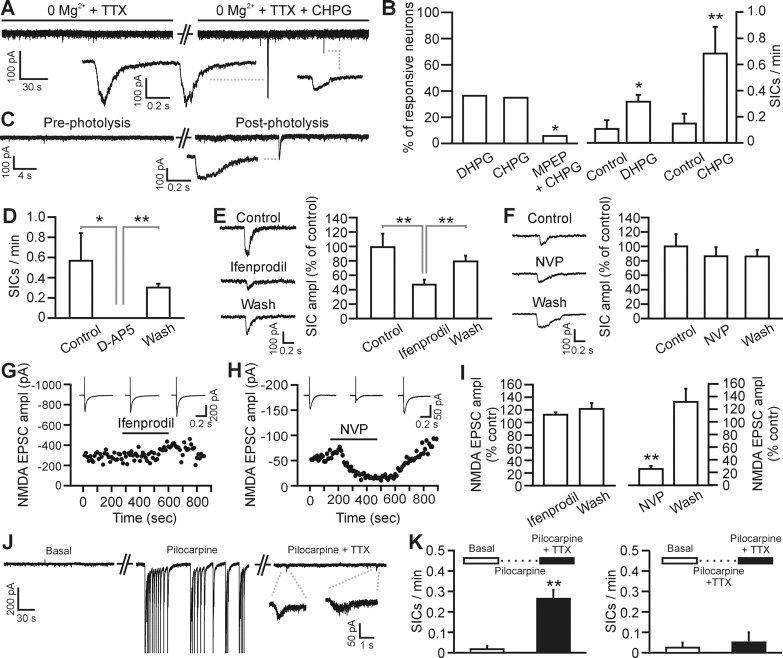
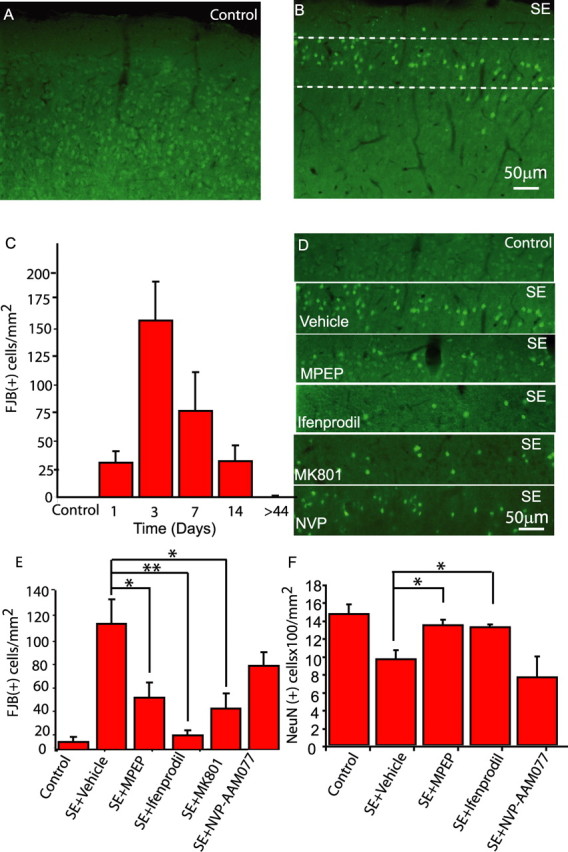
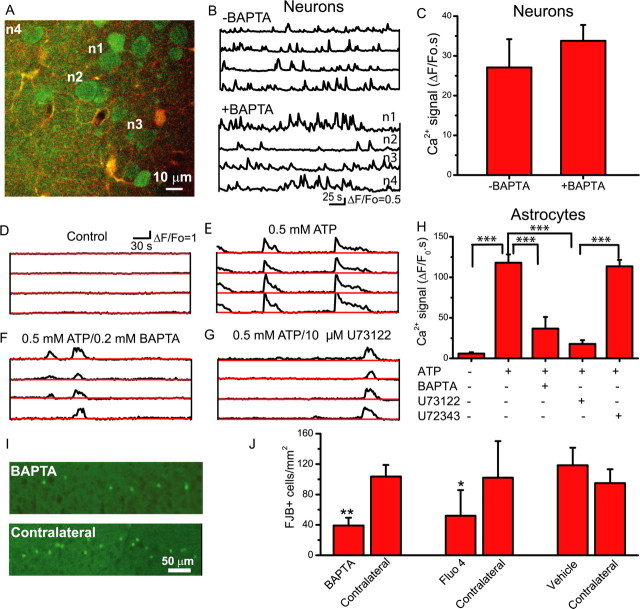
Status epilepticus (SE), an unremitting seizure, is known to cause a variety of traumatic responses including delayed neuronal death and later cognitive decline. Although excitotoxicity has been implicated in this delayed process, the cellular mechanisms are unclear. Because our previous brain slice studies have shown that chemically induced epileptiform activity can lead to elevated astrocytic Ca2+ signaling and because these signals are able to induce the release of the excitotoxic transmitter glutamate from these glia, we asked whether astrocytes are activated during status epilepticus and whether they contribute to delayed neuronal death in vivo. Using two-photon microscopy in vivo, we show that status epilepticus enhances astrocytic Ca2+ signals for 3 d and that the period of elevated glial Ca2+ signaling is correlated with the period of delayed neuronal death. To ask whether astrocytes contribute to delayed neuronal death, we first administered antagonists which inhibit gliotransmission: MPEP [2-methyl-6-(phenylethynyl)pyridine], a metabotropic glutamate receptor 5 antagonist that blocks astrocytic Ca2+ signals in vivo, and ifenprodil, an NMDA receptor antagonist that reduces the actions of glial-derived glutamate. Administration of these antagonists after SE provided significant neuronal protection raising the potential for a glial contribution to neuronal death. To test this glial hypothesis directly, we loaded Ca2+ chelators selectively into astrocytes after status epilepticus. We demonstrate that the selective attenuation of glial Ca2+ signals leads to neuronal protection. These observations support neurotoxic roles for astrocytic gliotransmission in pathological conditions and identify this process as a novel therapeutic target.
Figures

Similar articles
-
Time course and mechanism of hippocampal neuronal death in an in vitro model of status epilepticus: role of NMDA receptor activation and NMDA dependent calcium entry.Eur J Pharmacol. 2008 Mar 31;583(1):73-83. doi: 10.1016/j.ejphar.2008.01.025. Epub 2008 Feb 5. Eur J Pharmacol. 2008. PMID: 18289526 Free PMC article.
-
Photothrombosis ischemia stimulates a sustained astrocytic Ca2+ signaling in vivo.Glia. 2009 May;57(7):767-76. doi: 10.1002/glia.20804. Glia. 2009. PMID: 18985731 Free PMC article.
-
Mature hippocampal astrocytes exhibit functional metabotropic and ionotropic glutamate receptors in situ.Glia. 1999 Mar;26(1):1-11. doi: 10.1002/(sici)1098-1136(199903)26:1<1::aid-glia1>3.0.co;2-z. Glia. 1999. PMID: 10088667
-
Protective mechanisms of adenosine in neurons and glial cells.Ann N Y Acad Sci. 1997 Oct 15;825:1-10. doi: 10.1111/j.1749-6632.1997.tb48409.x. Ann N Y Acad Sci. 1997. PMID: 9369970 Review.
-
Bidirectional astrocyte-neuron communication: the many roles of glutamate and ATP.Novartis Found Symp. 2006;276:208-17; discussion 217-21, 233-7, 275-81. doi: 10.1002/9780470032244.ch16. Novartis Found Symp. 2006. PMID: 16805432 Review.
Cited by
-
Mitochondrial calcium cycling in neuronal function and neurodegeneration.Front Cell Dev Biol. 2023 Jan 24;11:1094356. doi: 10.3389/fcell.2023.1094356. eCollection 2023. Front Cell Dev Biol. 2023. PMID: 36760367 Free PMC article. Review.
-
Trafficking of immune cells in the central nervous system.J Clin Invest. 2010 May;120(5):1368-79. doi: 10.1172/JCI41911. Epub 2010 May 3. J Clin Invest. 2010. PMID: 20440079 Free PMC article. Review.
-
Astrocytic Ca2+ Signaling in Epilepsy.Front Cell Neurosci. 2021 Jul 15;15:695380. doi: 10.3389/fncel.2021.695380. eCollection 2021. Front Cell Neurosci. 2021. PMID: 34335188 Free PMC article. Review.
-
D-Serine Contributes to Seizure Development via ERK Signaling.Front Neurosci. 2019 Mar 26;13:254. doi: 10.3389/fnins.2019.00254. eCollection 2019. Front Neurosci. 2019. PMID: 30971878 Free PMC article.
-
Quantitative mass spectrometry-based proteomics reveals the dynamic range of primary mouse astrocyte protein secretion.J Proteome Res. 2010 May 7;9(5):2764-74. doi: 10.1021/pr100134n. J Proteome Res. 2010. PMID: 20329800 Free PMC article.
References
-
- Araque A, Parpura V, Sanzgiri RP, Haydon PG. Glutamate-dependent astrocyte modulation of synaptic transmission between cultured hippocampal neurons. Eur J Neurosci. 1998;10:2129–2142. - PubMed
-
- Araque A, Carmignoto G, Haydon PG. Dynamic signaling between astrocytes and neurons. Annu Rev Physiol. 2001;63:795–813. - PubMed
-
- Auberson YP, Allgeier H, Bischoff S, Lingenhoehl K, Moretti R, Schmutz M. 5-Phosphonomethylquinoxalinediones as competitive NMDA receptor antagonists with a preference for the human 1A/2A, rather than 1A/2B receptor composition. Bioorg Med Chem Lett. 2002;12:1099–1102. - PubMed
-
- Babb TL, Brown WJ, Pretorius J, Davenport C, Lieb JP, Crandall PH. Temporal lobe volumetric cell densities in temporal lobe epilepsy. Epilepsia. 1984;25:729–740. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous