

Caenorhabditis elegans prom-1 is required for meiotic prophase progression and homologous chromosome pairing
- PMID: 17914060
- PMCID: PMC2096575
- DOI: 10.1091/mbc.e07-03-0243
Caenorhabditis elegans prom-1 is required for meiotic prophase progression and homologous chromosome pairing
Abstract
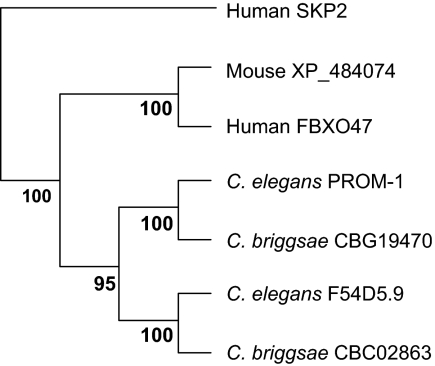
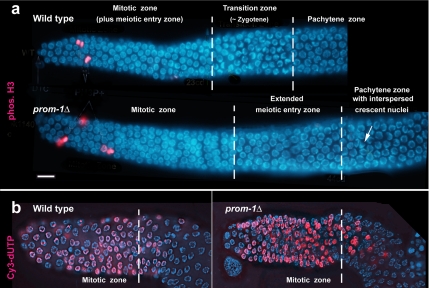
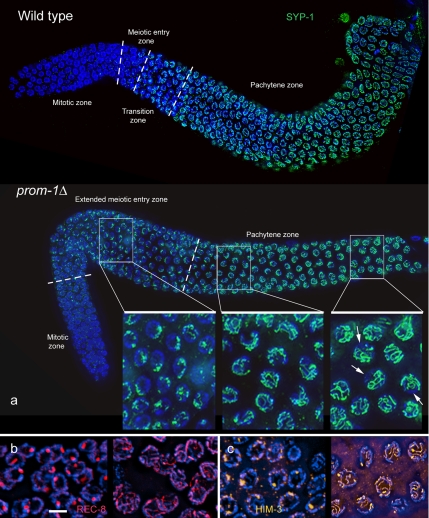
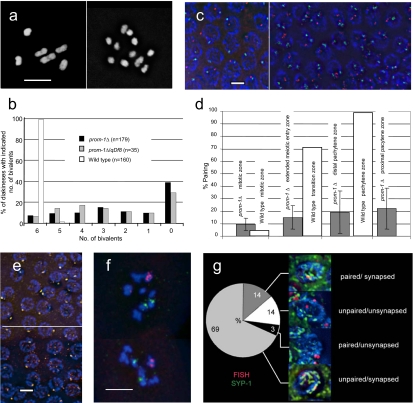
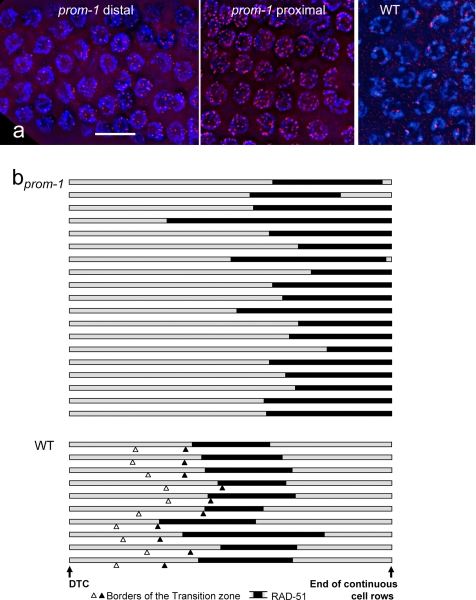
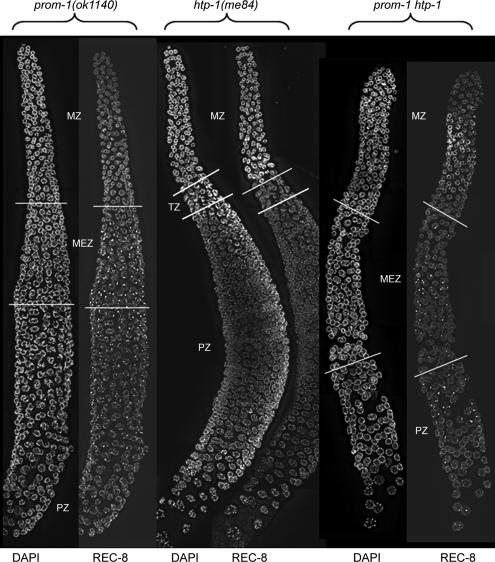
A novel gene, prom-1, was isolated in a screen for Caenorhabditis elegans mutants with increased apoptosis in the germline. prom-1 encodes an F-box protein with limited homology to the putative human tumor suppressor FBXO47. Mutations in the prom-1 locus cause a strong reduction in bivalent formation, which results in increased embryonic lethality and a Him phenotype. Furthermore, retarded and asynchronous nuclear reorganization as well as reduced homologous synapsis occur during meiotic prophase. Accumulation of recombination protein RAD-51 in meiotic nuclei suggests disturbed repair of double-stranded DNA breaks. Nuclei in prom-1 mutant gonads timely complete mitotic proliferation and premeiotic replication, but they undergo prolonged delay upon meiotic entry. We, therefore, propose that prom-1 regulates the timely progression through meiotic prophase I and that in its absence the recognition of homologous chromosomes is strongly impaired.
Figures
Similar articles
-
A quality control mechanism coordinates meiotic prophase events to promote crossover assurance.PLoS Genet. 2014 Apr 24;10(4):e1004291. doi: 10.1371/journal.pgen.1004291. eCollection 2014 Apr. PLoS Genet. 2014. PMID: 24762417 Free PMC article.
-
Caenorhabditis elegans HIM-18/SLX-4 interacts with SLX-1 and XPF-1 and maintains genomic integrity in the germline by processing recombination intermediates.PLoS Genet. 2009 Nov;5(11):e1000735. doi: 10.1371/journal.pgen.1000735. Epub 2009 Nov 20. PLoS Genet. 2009. PMID: 19936019 Free PMC article.
-
Synapsis-dependent and -independent mechanisms stabilize homolog pairing during meiotic prophase in C. elegans.Genes Dev. 2002 Sep 15;16(18):2428-42. doi: 10.1101/gad.1011602. Genes Dev. 2002. PMID: 12231631 Free PMC article.
-
Chromosome pairing and synapsis during Caenorhabditis elegans meiosis.Curr Opin Cell Biol. 2013 Jun;25(3):349-56. doi: 10.1016/j.ceb.2013.03.003. Epub 2013 Apr 8. Curr Opin Cell Biol. 2013. PMID: 23578368 Free PMC article. Review.
-
Homologue pairing, recombination and segregation in Caenorhabditis elegans.Genome Dyn. 2009;5:43-55. doi: 10.1159/000166618. Genome Dyn. 2009. PMID: 18948706 Review.
Cited by
-
Transient and Partial Nuclear Lamina Disruption Promotes Chromosome Movement in Early Meiotic Prophase.Dev Cell. 2018 Apr 23;45(2):212-225.e7. doi: 10.1016/j.devcel.2018.03.018. Dev Cell. 2018. PMID: 29689196 Free PMC article.
-
foxl3, a sexual switch in germ cells, initiates two independent molecular pathways for commitment to oogenesis in medaka.Proc Natl Acad Sci U S A. 2020 Jun 2;117(22):12174-12181. doi: 10.1073/pnas.1918556117. Epub 2020 May 14. Proc Natl Acad Sci U S A. 2020. PMID: 32409601 Free PMC article.
-
Release of CHK-2 from PPM-1.D anchorage schedules meiotic entry.Sci Adv. 2022 Feb 18;8(7):eabl8861. doi: 10.1126/sciadv.abl8861. Epub 2022 Feb 16. Sci Adv. 2022. PMID: 35171669 Free PMC article.
-
Mutations in Caenorhabditis elegans him-19 show meiotic defects that worsen with age.Mol Biol Cell. 2010 Mar 15;21(6):885-96. doi: 10.1091/mbc.e09-09-0811. Epub 2010 Jan 13. Mol Biol Cell. 2010. PMID: 20071466 Free PMC article.
-
Leptotene/zygotene chromosome movement via the SUN/KASH protein bridge in Caenorhabditis elegans.PLoS Genet. 2010 Nov 24;6(11):e1001219. doi: 10.1371/journal.pgen.1001219. PLoS Genet. 2010. PMID: 21124819 Free PMC article.
References
-
- Alpi A., Pasierbek P., Gartner A., Loidl J. Genetic and cytological characterization of the recombination protein RAD-51 in Caenorhabditis elegans. Chromosoma. 2003;112:6–16. - PubMed
-
- Baarends W. M., Wassenaar E., Hoogerbrugge J. W., van Cappellen G., Roest H. P., Vreeburg J., Ooms M., Hoeijmakers J.H.J., Grootegoed J. A. Loss of HR6B ubiquitin-conjugating activity results in damaged synaptonemal complex structure and increased crossing-over frequency during the male meiotic prophase. Mol. Cell Biol. 2003;23:1151–1162. - PMC - PubMed
-
- Bhalla N., Dernburg A. F. A conserved checkpoint monitors meiotic chromosome synapsis in Caenorhabditis elegans. Science. 2005;310:1683–1686. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
