

Dominant-negative behavior of mammalian Vps35 in yeast requires a conserved PRLYL motif involved in retromer assembly
- PMID: 17916227
- PMCID: PMC2532708
- DOI: 10.1111/j.1600-0854.2007.00658.x
Dominant-negative behavior of mammalian Vps35 in yeast requires a conserved PRLYL motif involved in retromer assembly
Abstract
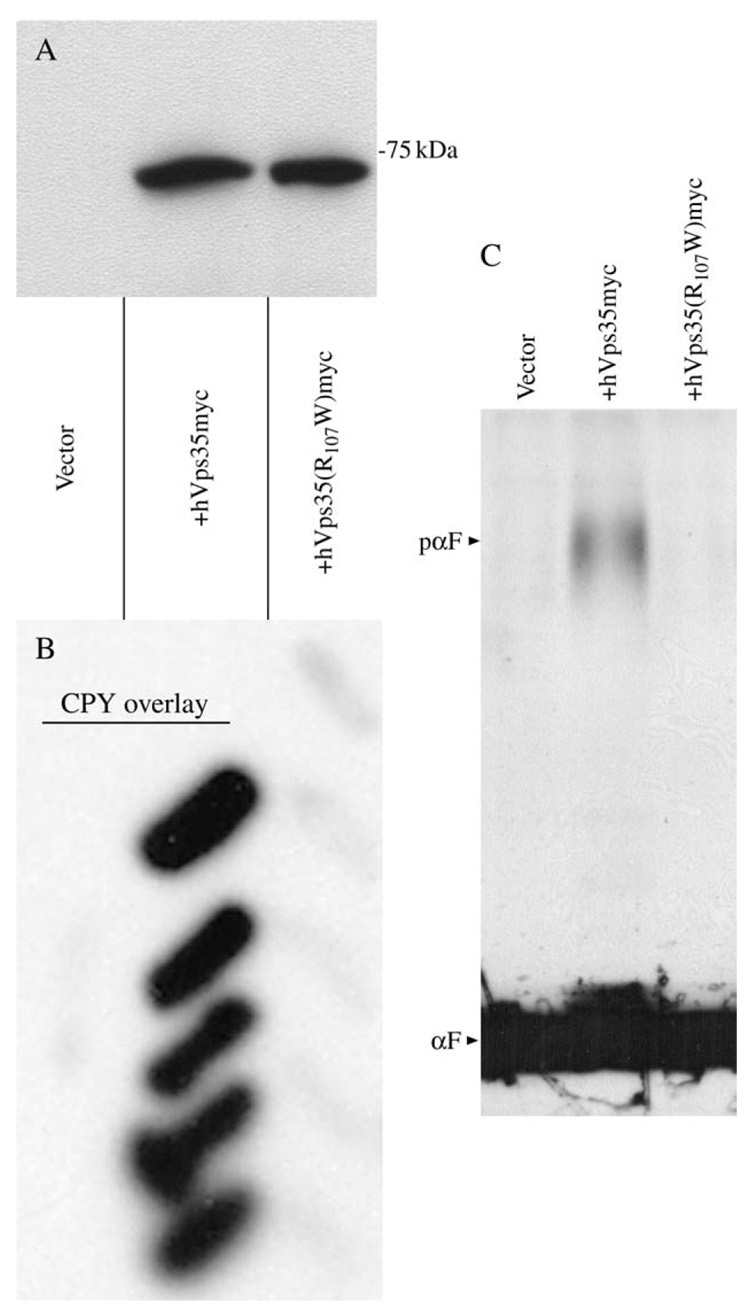
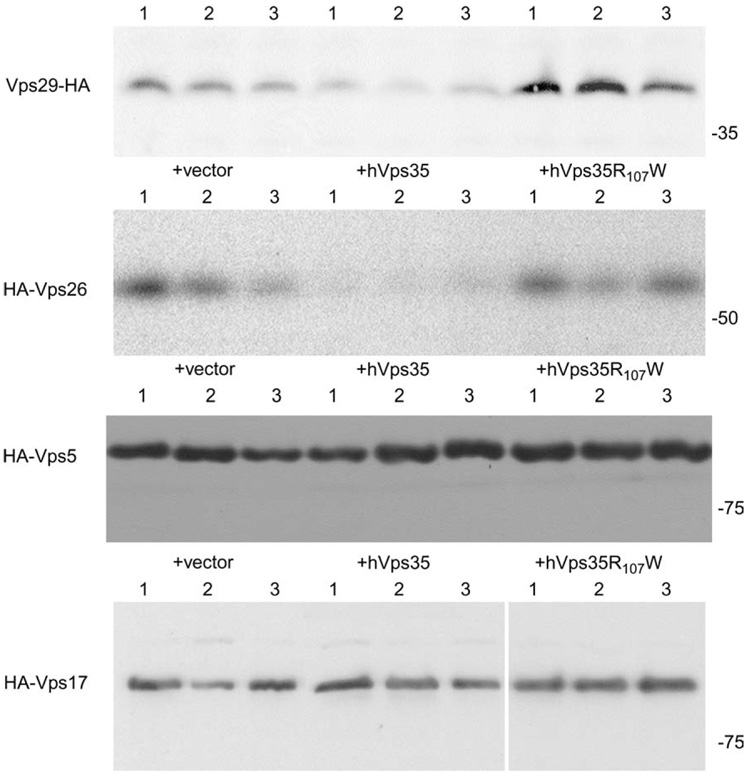
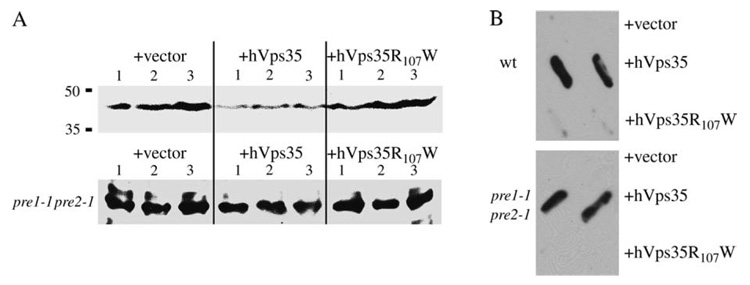
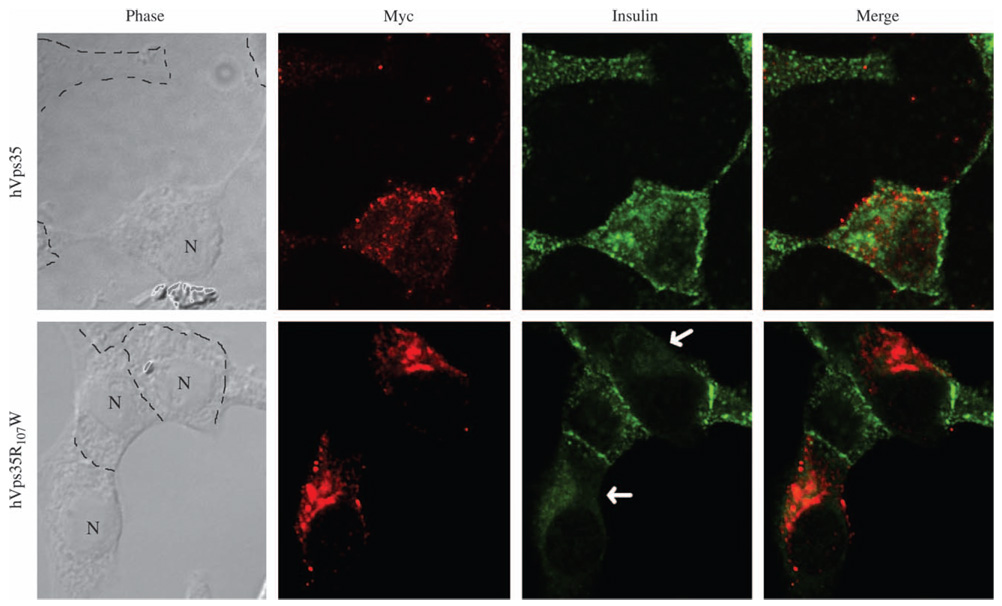
The retromer protein complex assists in recycling selected integral membrane proteins from endosomes to the trans Golgi network. One protein subcomplex (Vps35p, Vps26p and Vps29p) combines with a second (Vps17p and Vps5p) to form a coat involved in sorting and budding of endosomal vesicles. Yeast Vps35p (yVps35) exhibits similarity to human Vps35 (hVps35), especially in a completely conserved PRLYL motif contained within an amino-terminal domain. Companion studies indicate that an R(98)W mutation in yVps35 causes defective retromer assembly in Saccharomyces cerevisiae. Herein, we find that the expression of hVps35 in yeast confers dominant-negative vacuolar proenzyme secretion and defective secretory proprotein processing. The mutant phenotype appears to be driven by hVps35 competing with endogenous yVps35, becoming incorporated into defective retromer complexes and causing proteasomal degradation of endogenous Vps26 and Vps29. Increased expression of yVps35 displaces some hVps35 to a 100 000 x g supernatant and suppresses the dominant-negative phenotype. Remarkably, mutation of the conserved R(107)W of hVps35 displaces some of the protein to the 100 000 x g supernatant, slows protein turnover and restores stability of Vps26p and Vps29p and completely abrogates dominant-negative trafficking behavior. We show that hVps35 coprecipitates Vps26, whereas the R(107)W mutant does not. In pancreatic beta cells, the R(107)W mutant shifts hVps35 from peripheral endosomes to a juxtanuclear compartment, affecting both mannose phosphate receptors and insulin. These data underscore importance of the Vps35 PRLYL motif in retromer subcomplex interactions and function.
Figures







Similar articles
-
Structural features of vps35p involved in interaction with other subunits of the retromer complex.Traffic. 2007 Dec;8(12):1841-1853. doi: 10.1111/j.1600-0854.2007.00659.x. Epub 2007 Oct 17. Traffic. 2007. PMID: 17892535 Free PMC article.
-
Identification of a conserved motif required for Vps35p/Vps26p interaction and assembly of the retromer complex.Biochem J. 2007 Dec 1;408(2):287-95. doi: 10.1042/BJ20070555. Biochem J. 2007. PMID: 17696874 Free PMC article.
-
Human orthologs of yeast vacuolar protein sorting proteins Vps26, 29, and 35: assembly into multimeric complexes.Mol Biol Cell. 2000 Dec;11(12):4105-16. doi: 10.1091/mbc.11.12.4105. Mol Biol Cell. 2000. PMID: 11102511 Free PMC article.
-
Retromer: Structure, function, and roles in mammalian disease.Eur J Cell Biol. 2015 Nov;94(11):513-21. doi: 10.1016/j.ejcb.2015.07.002. Epub 2015 Jul 17. Eur J Cell Biol. 2015. PMID: 26220253 Review.
-
Unveiling the cryo-EM structure of retromer.Biochem Soc Trans. 2020 Oct 30;48(5):2261-2272. doi: 10.1042/BST20200552. Biochem Soc Trans. 2020. PMID: 33125482 Free PMC article. Review.
Cited by
-
The giardial VPS35 retromer subunit is necessary for multimeric complex assembly and interaction with the vacuolar protein sorting receptor.Biochim Biophys Acta. 2013 Dec;1833(12):2628-2638. doi: 10.1016/j.bbamcr.2013.06.015. Epub 2013 Jun 26. Biochim Biophys Acta. 2013. PMID: 23810936 Free PMC article.
-
EhVps35, a retromer component, is involved in the recycling of the EhADH and Gal/GalNac virulent proteins of Entamoeba histolytica.Front Parasitol. 2024 Mar 26;3:1356601. doi: 10.3389/fpara.2024.1356601. eCollection 2024. Front Parasitol. 2024. PMID: 39817169 Free PMC article.
-
Structural features of vps35p involved in interaction with other subunits of the retromer complex.Traffic. 2007 Dec;8(12):1841-1853. doi: 10.1111/j.1600-0854.2007.00659.x. Epub 2007 Oct 17. Traffic. 2007. PMID: 17892535 Free PMC article.
-
RME-8 coordinates the activity of the WASH complex with the function of the retromer SNX dimer to control endosomal tubulation.J Cell Sci. 2014 May 1;127(Pt 9):2053-70. doi: 10.1242/jcs.144659. Epub 2014 Mar 18. J Cell Sci. 2014. PMID: 24643499 Free PMC article.
-
Quantitative analysis of retromer complex-related genes during embryo development in the mouse.Mol Cells. 2011 May;31(5):431-6. doi: 10.1007/s10059-011-0272-7. Epub 2011 Feb 22. Mol Cells. 2011. PMID: 21359680 Free PMC article.
References
-
- Seaman MN. Recycle your receptors with retromer. Trends Cell Biol. 2005;15:68–75. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
