

Review
doi: 10.1016/j.sbi.2007.08.018.
Epub 2007 Oct 4.
Taking it step by step: mechanistic insights from structural studies of ubiquitin/ubiquitin-like protein modification pathways
Affiliations
- PMID: 17919899
- PMCID: PMC2174906
- DOI: 10.1016/j.sbi.2007.08.018
Item in Clipboard
Review
Taking it step by step: mechanistic insights from structural studies of ubiquitin/ubiquitin-like protein modification pathways
Curr Opin Struct Biol.
2007 Dec.
Abstract
Ubiquitin (Ub) and ubiquitin-like (Ubl) proteins regulate a diverse array of cellular pathways through post-translational attachment to protein substrates. Ub/Ubl-mediated signaling is initiated through E1, E2, and E3-mediated conjugation, transduced by proteins that recognize Ub/Ubl-modified substrates, and terminated by proteases which remove the Ub/Ubl from the substrate. Recent structural studies have elucidated mechanisms pertinent to Ub/Ubl conjugation, recognition, and deconjugation, highlighting essential steps during Ub/Ubl modification that illustrate common and divergent mechanistic themes within this important process.
Figures
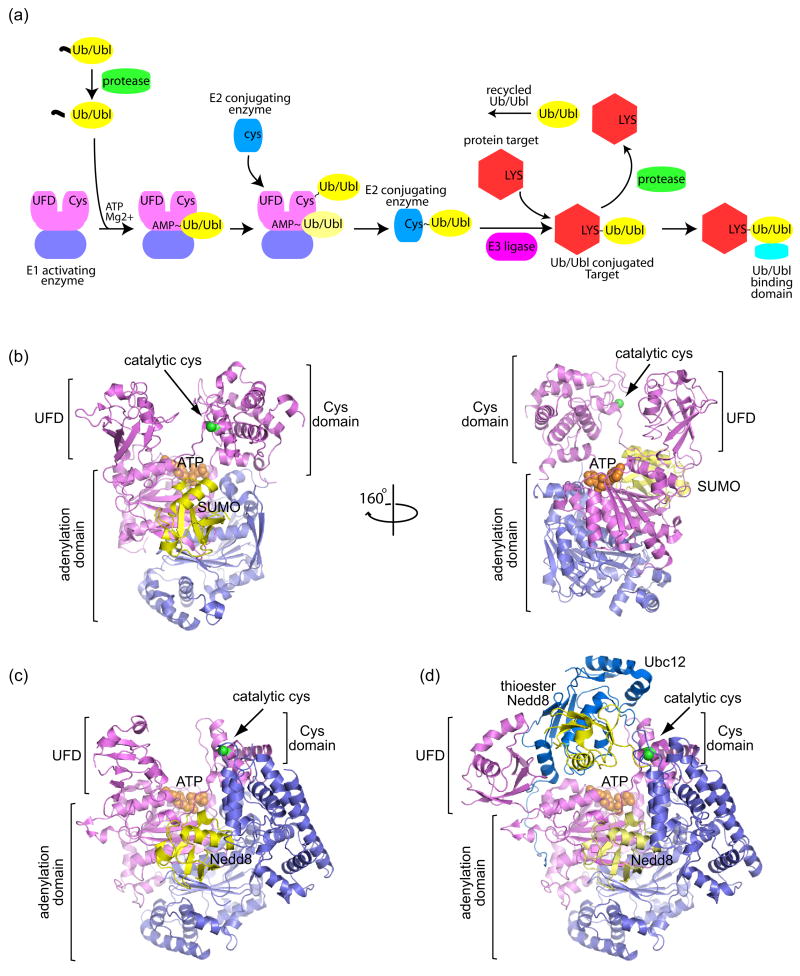
Schematic of the Ubiquitin/Ubiquitin-like (Ub/Ubl) protein modification pathway and structures of E1 activating enzymes. (a) The Ub/Ubl modification pathway begins with precursor Ub/Ubl (yellow) processing by proteases (green), exposing a C-terminal Gly-Gly motif. Ub/Ubl is activated by E1 enzymes (violet/pink) and transferred to a catalytic E1 cysteine residue to form a thioester bond (denoted by ‘~’). When the Ub/Ubl is linked to the E1 via a thioester, another Ub/Ubl may bind to the adenylation site (light yellow Ub/Ubl). E1 recruits a cognate E2 conjugating enzyme (blue) and the Ub/Ubl is transferred to the E2 via a thioester bond. Facilitated by an E3 ligase (magenta), the Ub/Ubl is conjugated to a Lys residue on a protein target (red), forming a stable isopeptide bond (denoted by ‘-‘). The Ub/Ubl conjugated protein target can be recognized by Ub/Ubl binding domains (cyan). The Ub/Ubl ‘signal’ can be terminated by deconjugating proteases. (b) Structure of the SUMO E1 Aos1/Uba2 (violet/pink) in complex with SUMO (yellow) and ATP (orange) (PDB code 1Y8R, [3]). The locations of the adenylation domain, Cys domain, and ubiquitin-fold domain (UFD) are indicated by labels and brackets. The SUMO E1 and Nedd8 E1 are heterodimeric. The adenylation domain is formed by both subunits, while the UFD and Cys domains are included within the Uba2 (for SUMO) and Uba3 (for Nedd8) subunits. ATP is shown in orange CPK within the adenylation active site. The catalytic cysteine on the Cys domain is denoted by a green sphere. The right panel shows the SUMO E1 rotated by 160° to illustrate the large distance between the ATP and E1 catalytic cysteine. (c) Structure of Nedd8 E1 APPBP1/Uba3 (violet/pink) in complex with Nedd8 (yellow) and ATP (orange) (PDB code 1R4N, [5]). (d) Structure of the double-loaded Nedd8~E1-Nedd8-E2 in which Nedd8 is captured in the adenylation site and as a thioester linked adduct to the E1 Cys domain (PDB code 2NVU, [6••]). The Nedd8 E2, Ubc12 (blue), is recruited by the UFD domain and interacts with the UFD, adenylation, and thioester linked Nedd8. The UFD is rotated 120° from that in (c) to accommodate the E2. All structural representations created with PyMOL (http://pymol.sourceforge.net/ ).
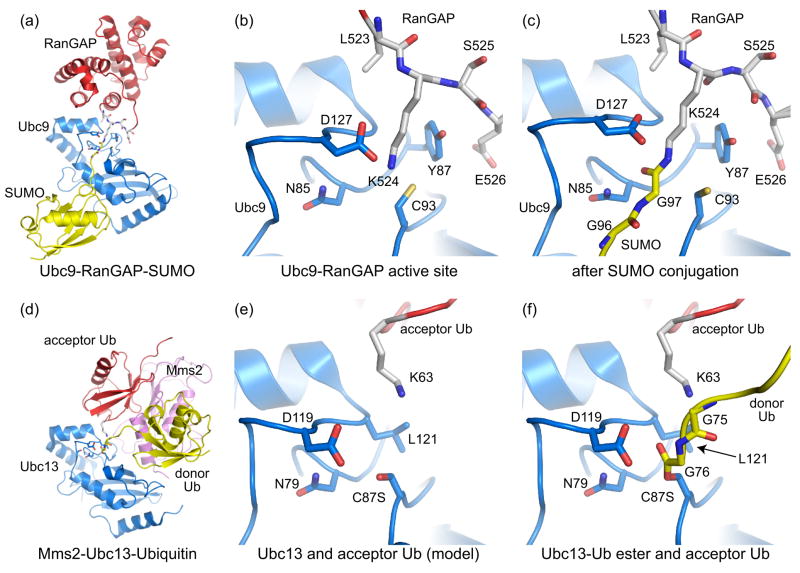
Recognition of substrate acceptor lysine residues by E2 conjugating enzymes. (a) Structure depicting Ubc9 (blue) in complex with RanGAP (red) conjugated to SUMO (yellow) (PDB code 1Z5S, [11•]). Nup358 is not shown. (b) Ubc9-RanGAP active site in a pre-conjugation state (PDB code 2GRN, [12••]). (c) SUMO-conjugated RanGAP-Ubc9 complex showing the isopeptide bond and E2 active site in a post-conjugation state [11•]. Active site residues described in the text are shown. (d) Mms2-Ubc13-Ubiquitin complex containing Ubc13 (blue), Mms2 (pink), and ubiquitin (yellow) ester-linked to Ubc13 (PDB code 2GMI, [14••]). The donor Ub contacted a symmetry related Mms2, inserting its K63 residue into the active site of the symmetry related Ubc13 partner. Therefore, the acceptor Ub (red) originates from a symmetry related complex. (e) Ubc13 active site with the ester-linked Ub removed to facilitate comparisons between the E2 active sites and lysine positions in (b). (f) The Ubc13 active site indicating ester-linked donor Ub (yellow) and acceptor Ub (red).
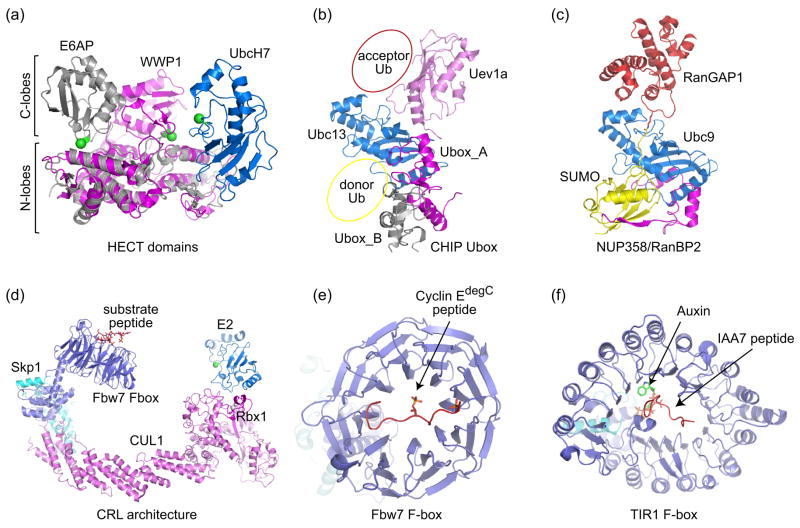
E3 Ub/Ubl ligases. (a) Structure of the HECT domain E6AP (grey) in complex with the E2 UbcH7 (blue) (PDB code 1C4Z, [20]). The HECT domain WWP1 (magenta) (PDB code 1ND7, [21]) is superimposed onto the E6AP HECT domain via their N-terminal lobes to demonstrate the relative movement of the C-terminal lobe towards the E2. The N- and C-lobes are indicated by labels and brackets. The catalytic cysteines for both HECT domains and UbcH7 are denoted by green spheres. (b) CHIP U-box homodimer (grey and magenta) bound to Ubc13/Uev1a (blue/pink) (PDB code 2C2V, [18]). Donor Ub and acceptor Ub positions are suggested based on Ubc1~Ub [19], SUMO-RanGAP-Ubc9 [11•], and Mms2-Ubc13-Ub structures [14••]. (c) Complex between SUMO-conjugated-RanGAP, the SUMO E2 Ubc9, and an E3 Nup358 (magenta) (PDB code 1Z5S, [11•]). Contacts between SUMO and N-terminal IR1 are mediated by an anti-parallel β-strand that contacts an exposed SUMO β-sheet and α-helix, an interaction now recognized as a common feature in proteins that contain SUMO interaction motifs (see Figure 4c and 4d). (d) Model of a Cullin-RING ligase (CRL) illustrating positions for the E2 and substrate receptors. The model was constructed by docking the F-box Fbw7 (PDB code 2OVQ, [31]) complex onto the structure of Cul1-Rbx1-Skp1 SCF (PDB code 1LDK, [29]). The E2 (catalytic cysteine denoted by a green sphere) is modeled based on the Cbl-RING-UbcH7 structure (PDB code 1FBV, [50]). (e) Structure of the Fbw7 F-box bound to phosphorylated cyclin E C-terminal degron [31]. (f) TIR1 F-box in complex with Auxin and IAA7 peptide (PDB code 2P1Q, [32••]).
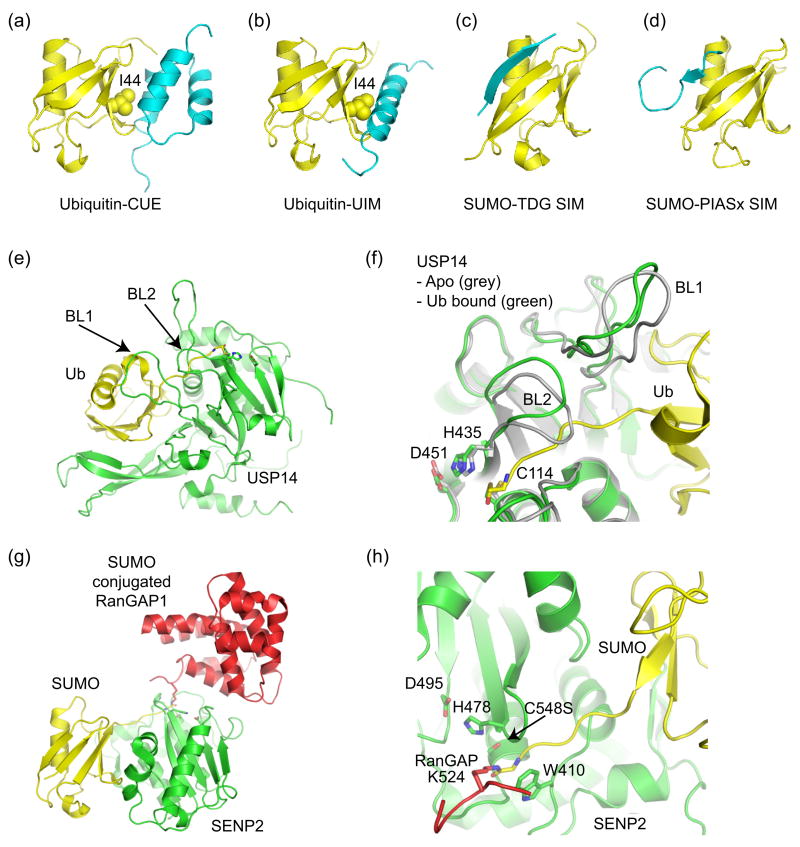
Ub/Ubl binding domains and Ub/Ubl proteases. (a) Ub in complex with CUE (PDB code 1OTR, [36,37]). (b) Ub in complex with UIM (PDB code 1Q0W, [35]). CUE and UIM domains recognize the Ub hydrophobic patch centered about Ile44. (c) SIM from thymine DNA glycosylase (TDG) in complex with SUMO indicating an anti-parallel orientation for the SIM β-strand (PDB code 1WYW, [39]), similar to that observed for the Nup358 IR1 SIM domain (Figure 3c). (d) SIM from PIASx adopts a parallel β-strand orientation in complex with SUMO (PDB code 2ASQ, [40]). (e) Structure of Ub protease USP14 in complex with Ub-aldehyde (PDB code 2AYO, [43]). Blocking loops (BL1 and BL2) are denoted. (f) In Apo-USP14 (grey) (PDB code 2AYN), BL1 and BL2 occlude access to the preformed catalytic site. In USP14-Ub-aldehyde (green), Ub induces a conformational change wherein BL1 and BL2 move away to provide access to the catalytic site. Active site residues are shown. (g) Structure of SUMO protease SENP2 in complex with SUMO conjugated RanGAP (PDB code 2IO2, [47•]). (h) Close-up of the SENP2 active site to illustrate the 90° kink and isopeptide bond between RanGAP and SUMO. Similar structures were observed for SENP1 [48•]. Active site residues are shown.
References
-
- Kerscher O, Felberbaum R, Hochstrasser M. Modification of proteins by ubiquitin and ubiquitin-like proteins. Annu Rev Cell Dev Biol. 2006;22:159–180. - PubMed
-
- Hershko A, Ciechanover A. The ubiquitin system. Annu Rev Biochem. 1998;67:425–479. - PubMed
-
- Walden H, Podgorski MS, Schulman BA. Insights into the ubiquitin transfer cascade from the structure of the activating enzyme for NEDD8. Nature. 2003;422:330–334. - PubMed
-
- Walden H, Podgorski MS, Huang DT, Miller DW, Howard RJ, Minor DL, Jr, Holton JM, Schulman BA. The structure of the APPBP1-UBA3-NEDD8-ATP complex reveals the basis for selective ubiquitin-like protein activation by an E1. Mol Cell. 2003;12:1427–1437. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
