

Drosophila Spd-2 recruits PCM to the sperm centriole, but is dispensable for centriole duplication
- PMID: 17919907
- PMCID: PMC2045633
- DOI: 10.1016/j.cub.2007.08.065
Drosophila Spd-2 recruits PCM to the sperm centriole, but is dispensable for centriole duplication
Abstract
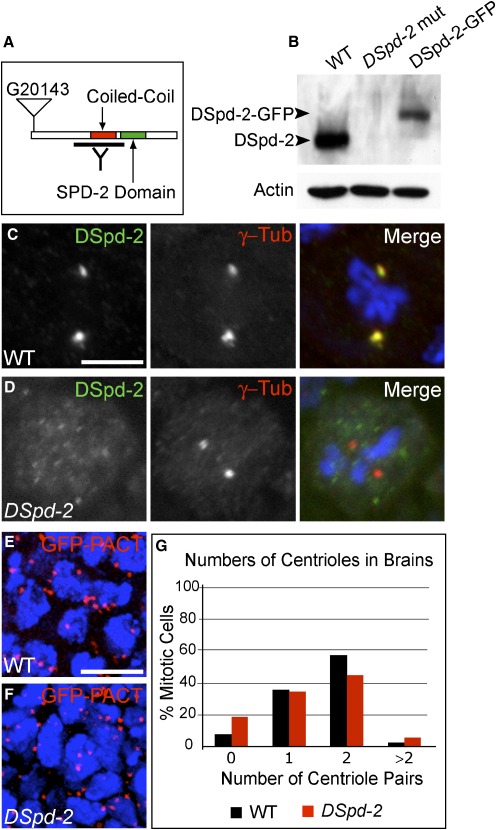
In C. elegans, genome-wide screens have identified just five essential centriole-duplication factors: SPD-2, ZYG-1, SAS-5, SAS-6, and SAS-4 [1-8]. These proteins are widely believed to comprise a conserved core duplication module [3, 9-14]. In worm embryos, SPD-2 is the most upstream component of this module, and it is also essential for pericentriolar material (PCM) recruitment to the centrioles [1, 4, 15, 16]. Here, we show that Drosophila Spd-2 (DSpd-2) is a component of both the centrioles and the PCM and has a role in recruiting PCM to the centrioles. DSpd-2 appears not, however, to be essential for centriole duplication in somatic cells. Moreover, PCM recruitment in DSpd-2 mutant somatic cells is only partially compromised, and mitosis appears unperturbed. In contrast, DSpd-2 is essential for proper PCM recruitment to the fertilizing sperm centriole, and hence for microtubule nucleation and pronuclear fusion. DSpd-2 therefore appears to have a particularly important role in recruiting PCM to the sperm centriole. We speculate that the SPD-2 family of proteins might only be absolutely essential for the recruitment of centriole duplication factors and PCM to the centriole(s) that enter the egg with the fertilizing sperm.
Figures



References
-
- Pelletier L., Ozlu N., Hannak E., Cowan C., Habermann B., Ruer M., Muller-Reichert T., Hyman A.A. The Caenorhabditis elegans centrosomal protein SPD-2 is required for both pericentriolar material recruitment and centriole duplication. Curr. Biol. 2004;14:863–873. - PubMed
-
- Leidel S., Gonczy P. SAS-4 is essential for centrosome duplication in C elegans and is recruited to daughter centrioles once per cell cycle. Dev. Cell. 2003;4:431–439. - PubMed
-
- Leidel S., Delattre M., Cerutti L., Baumer K., Gonczy P. SAS-6 defines a protein family required for centrosome duplication in C. elegans and in human cells. Nat. Cell Biol. 2005;7:115–125. - PubMed
-
- Kemp C.A., Kopish K.R., Zipperlen P., Ahringer J., O'Connell K.F. Centrosome maturation and duplication in C. elegans require the coiled-coil protein SPD-2. Dev. Cell. 2004;6:511–523. - PubMed
-
- Dammermann A., Muller-Reichert T., Pelletier L., Habermann B., Desai A., Oegema K. Centriole assembly requires both centriolar and pericentriolar material proteins. Dev. Cell. 2004;7:815–829. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
