

Exploring the parameter space of complex self-assembly through virus capsid models
- PMID: 17921216
- PMCID: PMC2186238
- DOI: 10.1529/biophysj.107.107284
Exploring the parameter space of complex self-assembly through virus capsid models
Abstract
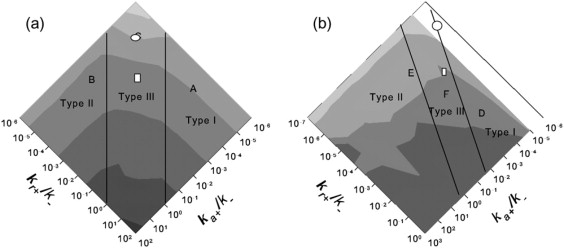
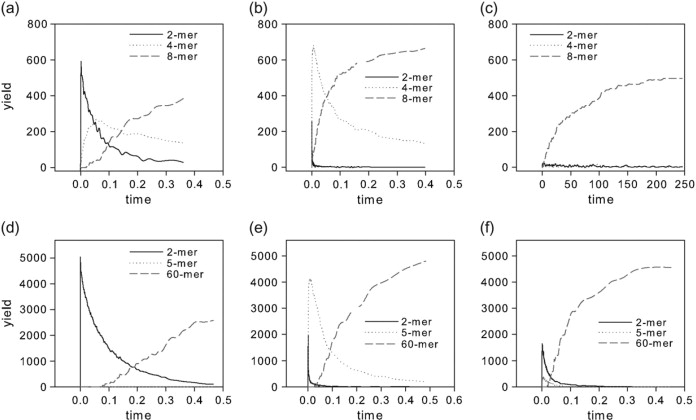
We use discrete event stochastic simulations to characterize the parameter space of a model of icosahedral viral capsid assembly as functions of monomer-monomer binding rates. The simulations reveal a parameter space characterized by three major assembly mechanisms, a standard nucleation-limited monomer-accretion pathway and two distinct hierarchical assembly pathways, as well as unproductive regions characterized by kinetically trapped species. Much of the productive parameter space also consists of border regions between these domains where hybrid pathways are likely to operate. A simpler octamer system studied for comparison reveals three analogous pathways, but is characterized by much lesser sensitivity to parameter variations in contrast to the sharp changes visible in the icosahedral model. The model suggests that modest changes in assembly conditions, consistent with expected differences between in vitro and in vivo assembly environments, could produce substantial shifts in assembly pathways. These results suggest that we must be cautious in drawing conclusions about in vivo capsid self-assembly dynamics from theoretical or in vitro models, as the nature of the basic assembly mechanisms accessible to a system can substantially differ between simple and complex model systems, between theoretical models and simulation results, and between in vitro and in vivo assembly conditions.
Figures
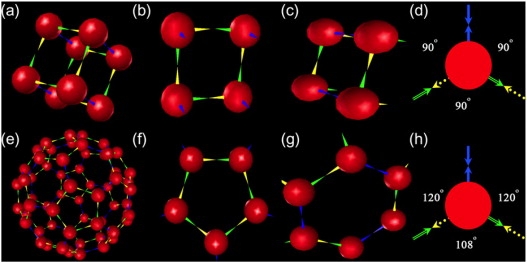
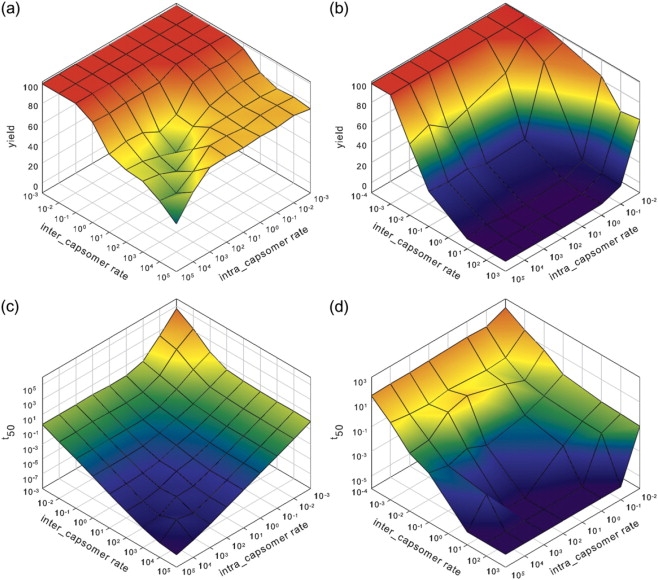
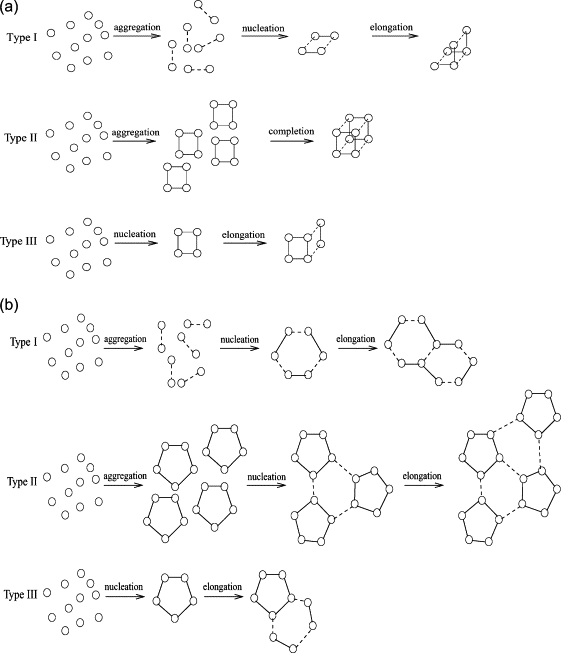
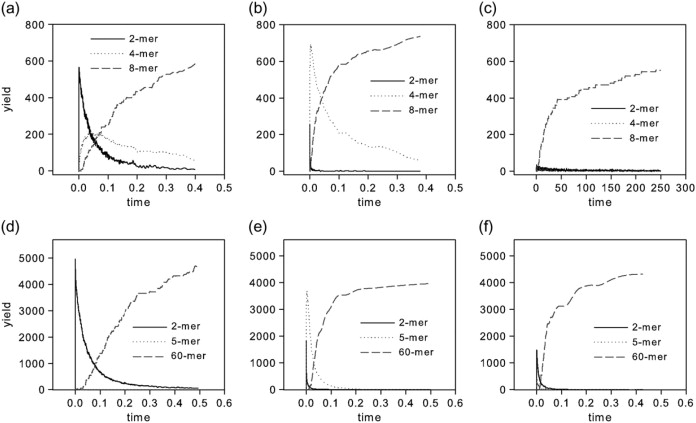
References
-
- Baskakov I.V., Aagaard C., Mehlhorn I., Wille H., Groth D., Baldwin M.A., Prusiner S.B., Cohen F.E. Self-assembly of recombinant prion protein of 106 residues. Biochemistry. 2000;39:2792–2804. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
