

Variant estrogen receptor-c-Src molecular interdependence and c-Src structural requirements for endothelial NO synthase activation
- PMID: 17921256
- PMCID: PMC2034248
- DOI: 10.1073/pnas.0704315104
Variant estrogen receptor-c-Src molecular interdependence and c-Src structural requirements for endothelial NO synthase activation
Abstract
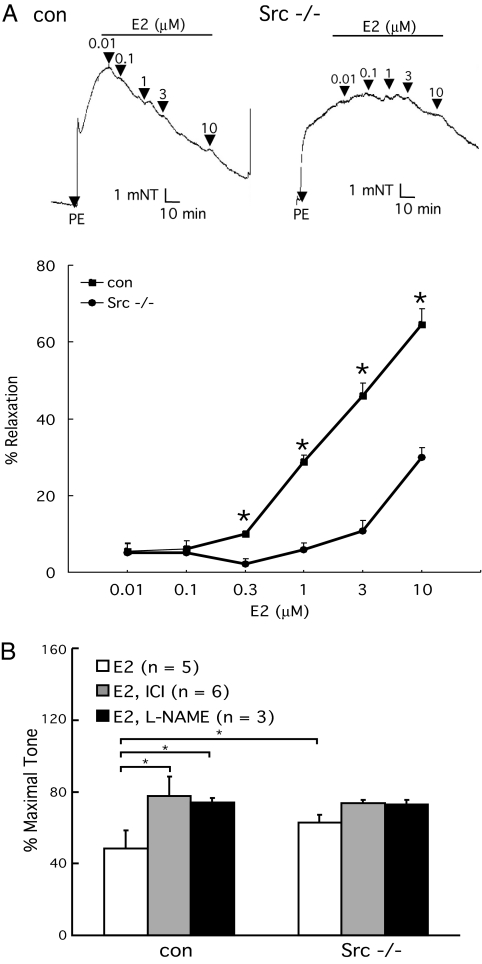
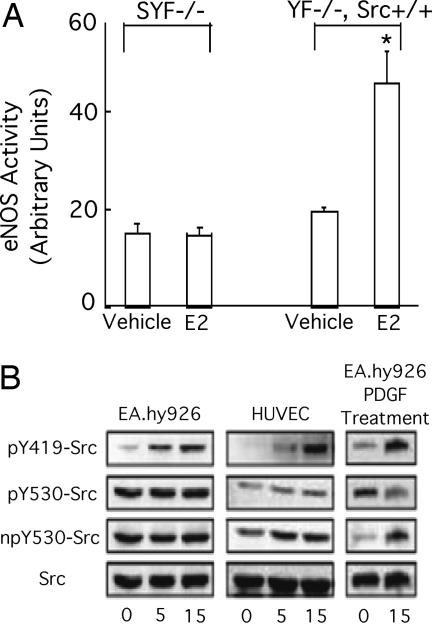
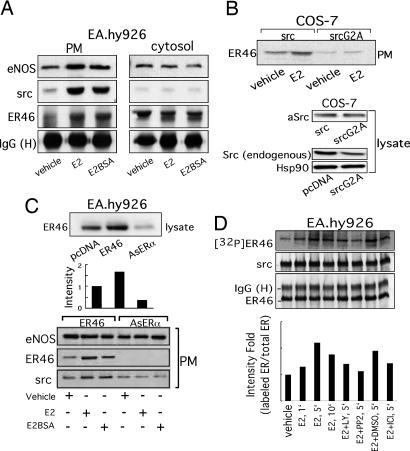
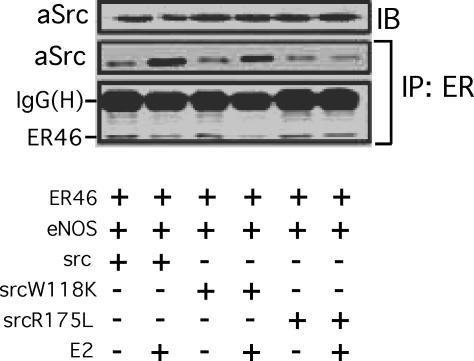
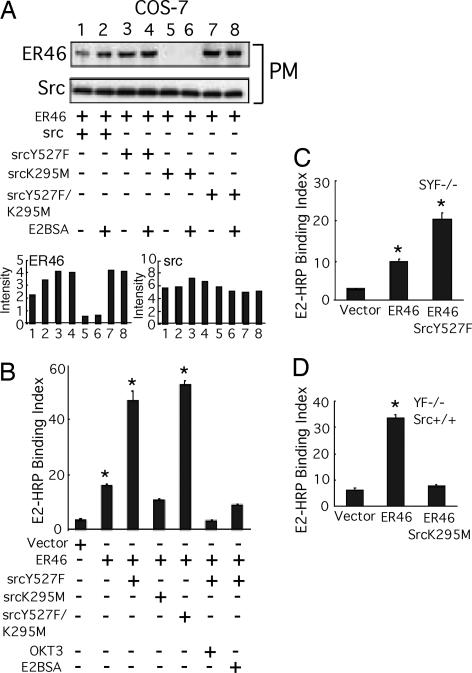
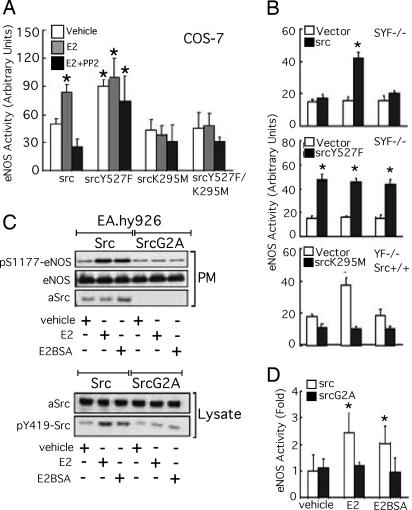
Little is known about the tyrosine kinase c-Src's function in the systemic circulation, in particular its role in arterial responses to hormonal stimuli. In human aortic and venous endothelial cells, c-Src is indispensable for 17beta-estradiol (E2)-stimulated phosphatidylinositol 3-kinase/Akt/endothelial NO synthase (eNOS) pathway activation, a possible mechanism in E2-mediated vascular protection. Here we show that c-Src supports basal and E2-stimulated NO production and is required for E2-induced vasorelaxation in murine aortas. Only membrane c-Src is structurally and functionally involved in E2-induced eNOS activation. Independent of c-Src kinase activity, c-Src is associated with an N-terminally truncated estrogen receptor alpha variant (ER46) and eNOS in the plasma membrane through its "open" (substrate-accessible) conformation. In the presence of E2, c-Src kinase is activated by membrane ER46 and in turn phosphorylates ER46 for subsequent ER46 and c-Src membrane recruitment, the assembly of an eNOS-centered membrane macrocomplex, and membrane-initiated eNOS activation. Overall, these results provide insights into a critical role for the tyrosine kinase c-Src in estrogen-stimulated arterial responses, and in membrane-initiated rapid signal transduction, for which obligate complex assembly and localization require the c-Src substrate-accessible structure.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
