

Two calcium-dependent protein kinases, CPK4 and CPK11, regulate abscisic acid signal transduction in Arabidopsis
- PMID: 17921317
- PMCID: PMC2174700
- DOI: 10.1105/tpc.107.050666
Two calcium-dependent protein kinases, CPK4 and CPK11, regulate abscisic acid signal transduction in Arabidopsis
Abstract
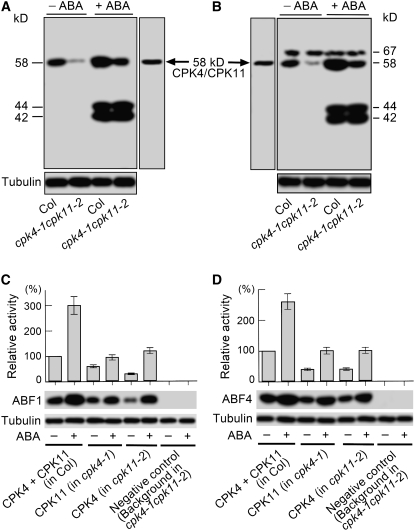
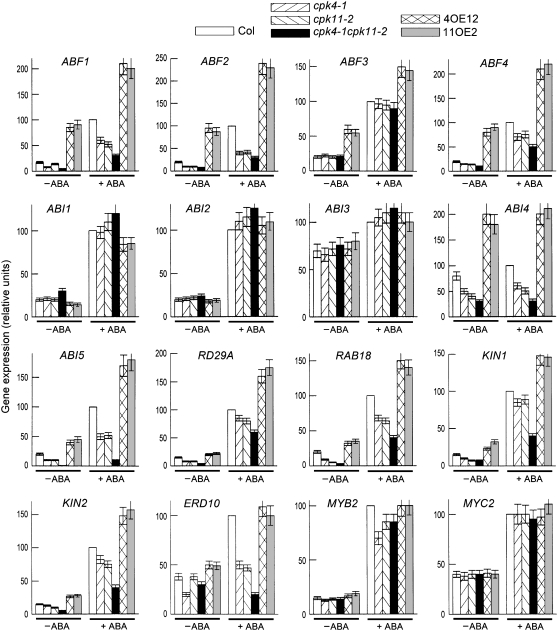
Many biochemical approaches show functions of calcium-dependent protein kinases (CDPKs) in abscisic acid (ABA) signal transduction, but molecular genetic evidence linking defined CDPK genes with ABA-regulated biological functions at the whole-plant level has been lacking. Here, we report that ABA stimulated two homologous CDPKs in Arabidopsis thaliana, CPK4 and CPK11. Loss-of-function mutations of CPK4 and CPK11 resulted in pleiotropic ABA-insensitive phenotypes in seed germination, seedling growth, and stomatal movement and led to salt insensitivity in seed germination and decreased tolerance of seedlings to salt stress. Double mutants of the two CDPK genes had stronger ABA- and salt-responsive phenotypes than the single mutants. CPK4- or CPK11-overexpressing plants generally showed inverse ABA-related phenotypes relative to those of the loss-of-function mutants. Expression levels of many ABA-responsive genes were altered in the loss-of-function mutants and overexpression lines. The CPK4 and CPK11 kinases both phosphorylated two ABA-responsive transcription factors, ABF1 and ABF4, in vitro, suggesting that the two kinases may regulate ABA signaling through these transcription factors. These data provide in planta genetic evidence for the involvement of CDPK/calcium in ABA signaling at the whole-plant level and show that CPK4 and CPK11 are two important positive regulators in CDPK/calcium-mediated ABA signaling pathways.
Figures

Similar articles
-
The Arabidopsis Ca(2+) -dependent protein kinase CPK12 negatively regulates abscisic acid signaling in seed germination and post-germination growth.New Phytol. 2011 Oct;192(1):61-73. doi: 10.1111/j.1469-8137.2011.03793.x. Epub 2011 Jun 21. New Phytol. 2011. PMID: 21692804
-
Abscisic acid inhibits root growth in Arabidopsis through ethylene biosynthesis.Plant J. 2014 Jul;79(1):44-55. doi: 10.1111/tpj.12534. Epub 2014 May 30. Plant J. 2014. PMID: 24738778
-
Expression of a grape calcium-dependent protein kinase ACPK1 in Arabidopsis thaliana promotes plant growth and confers abscisic acid-hypersensitivity in germination, postgermination growth, and stomatal movement.Plant Mol Biol. 2007 Jul;64(5):531-8. doi: 10.1007/s11103-007-9172-9. Epub 2007 May 3. Plant Mol Biol. 2007. PMID: 17476573
-
Abscisic acid signaling in seeds and seedlings.Plant Cell. 2002;14 Suppl(Suppl):S15-45. doi: 10.1105/tpc.010441. Plant Cell. 2002. PMID: 12045268 Free PMC article. Review. No abstract available.
-
Seeing 'cool' and 'hot'--infrared thermography as a tool for non-invasive, high-throughput screening of Arabidopsis guard cell signalling mutants.J Exp Bot. 2004 May;55(400):1187-93. doi: 10.1093/jxb/erh135. Epub 2004 Apr 8. J Exp Bot. 2004. PMID: 15073209 Review.
Cited by
-
Calcium-dependent protein kinase 2 plays a positive role in the salt stress response in potato.Plant Cell Rep. 2022 Mar;41(3):535-548. doi: 10.1007/s00299-021-02676-7. Epub 2021 Mar 2. Plant Cell Rep. 2022. PMID: 33651205
-
Transcription factors involved in drought tolerance and their possible role in developing drought tolerant cultivars with emphasis on wheat (Triticum aestivum L.).Theor Appl Genet. 2016 Nov;129(11):2019-2042. doi: 10.1007/s00122-016-2794-z. Epub 2016 Oct 13. Theor Appl Genet. 2016. PMID: 27738714 Review.
-
Interaction between Calcium and Actin in Guard Cell and Pollen Signaling Networks.Plants (Basel). 2013 Oct 15;2(4):615-34. doi: 10.3390/plants2040615. Plants (Basel). 2013. PMID: 27137395 Free PMC article. Review.
-
Genome-wide identification, and characterization of the CDPK gene family reveal their involvement in abiotic stress response in Fragaria x ananassa.Sci Rep. 2020 Jul 6;10(1):11040. doi: 10.1038/s41598-020-67957-9. Sci Rep. 2020. PMID: 32632235 Free PMC article.
-
Calcium-dependent protein kinases responsible for the phosphorylation of a bZIP transcription factor FD crucial for the florigen complex formation.Sci Rep. 2015 Feb 9;5:8341. doi: 10.1038/srep08341. Sci Rep. 2015. PMID: 25661797 Free PMC article.
References
-
- Achard, P., Cheng, H., Grauve, L.D., Decat, J., Schoutteten, H., Moritz, T., Straeten, V.D., Peng, J., and Harberd, N.P. (2006). Integration of plant responses to environmentally activated phytohormonal signals. Science 311 91–94. - PubMed
-
- Bradford, M.M. (1976). A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 72 248–254. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
