

Development and function of murine B220+CD11c+NK1.1+ cells identify them as a subset of NK cells
- PMID: 17923504
- PMCID: PMC2118497
- DOI: 10.1084/jem.20070991
Development and function of murine B220+CD11c+NK1.1+ cells identify them as a subset of NK cells
Abstract
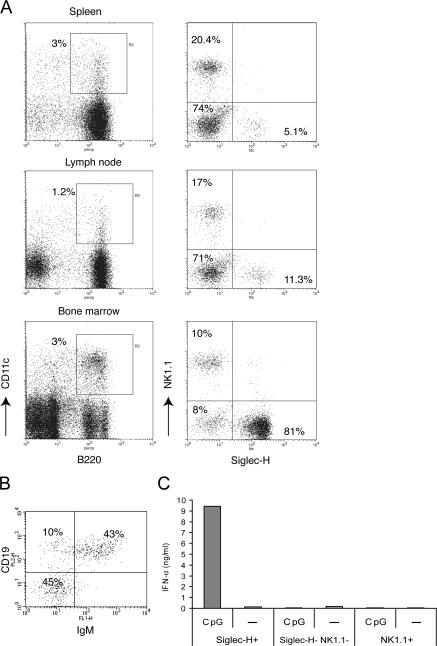
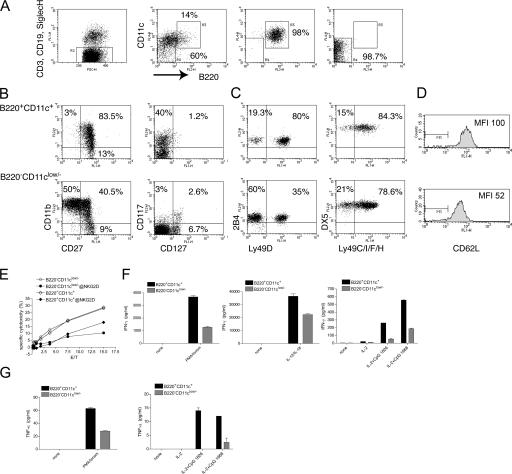
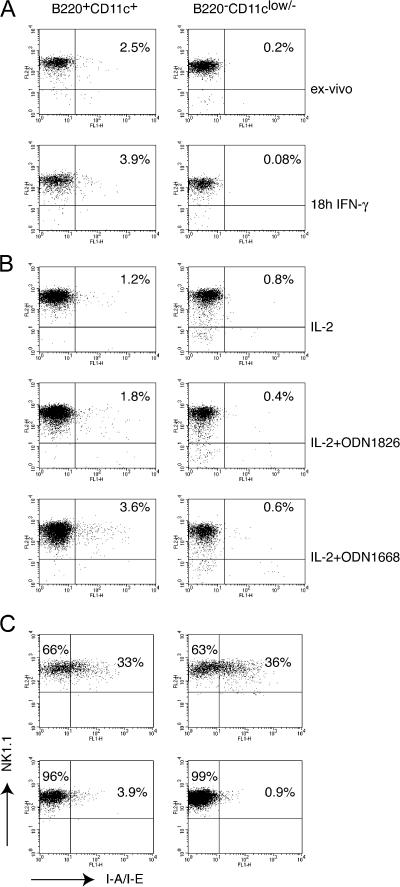
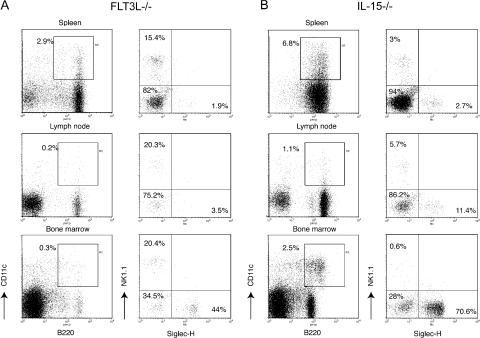
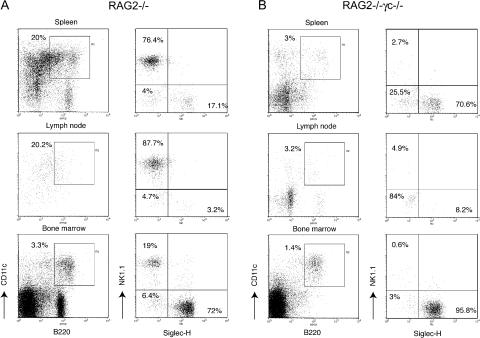
Lymphoid organs contain a B220(+)CD11c(+)NK1.1(+) cell population that was recently characterized as a novel dendritic cell (DC) subset that functionally overlaps with natural killer (NK) cells and plasmacytoid DCs (PDCs). Using Siglec-H and NK1.1 markers, we unambiguously dissected B220(+)CD11c(+) cells and found that PDCs are the only professional interferon (IFN)-alpha-producing cells within this heterogeneous population. In contrast, B220(+)CD11c(+)NK1.1(+) cells are a discrete NK cell subset capable of producing higher levels of IFN-gamma than conventional NK cells. Unlike DCs, only a minute fraction of B220(+)CD11c(+)NK1.1(+) cells in the spleen expressed major histocompatibility complex class II ex vivo or after stimulation with CpG. Consistent with being a NK cell subset, B220(+)CD11c(+)NK1.1(+) cells depended primarily on interleukin 15 and common cytokine receptor gamma chain signaling for their development. In terms of function, expression of distinctive cell surface receptors, and location in lymphoid organs, NK1.1(+)B220(+)CD11c(+) appear to be the murine equivalent of human CD56(bright) NK cells.
Figures
References
-
- Asselin-Paturel, C., A. Boonstra, M. Dalod, I. Durand, N. Yessaad, C. Dezutter-Dambuyant, A. Vicari, A. O'Garra, C. Biron, F. Briere, and G. Trinchieri. 2001. Mouse type I IFN-producing cells are immature APCs with plasmacytoid morphology. Nat. Immunol. 2:1144–1150. - PubMed
-
- Taieb, J., N. Chaput, C. Menard, L. Apetoh, E. Ullrich, M. Bonmort, M. Pequignot, N. Casares, M. Terme, C. Flament, et al. 2006. A novel dendritic cell subset involved in tumor immunosurveillance. Nat. Med. 12:214–219. - PubMed
-
- Chan, C.W., E. Crafton, H.N. Fan, J. Flook, K. Yoshimura, M. Skarica, D. Brockstedt, T.W. Dubensky, M.F. Stins, L.L. Lanier, et al. 2006. Interferon-producing killer dendritic cells provide a link between innate and adaptive immunity. Nat. Med. 12:207–213. - PubMed
-
- Vremec, D., M. O'Keeffe, H. Hochrein, M. Fuchsberger, I. Caminschi, M. Lahoud, and K. Shortman. 2007. Production of interferons by dendritic cells, plasmacytoid cells, natural killer cells, and interferon-producing killer dendritic cells. Blood. 109:1165–1173. - PubMed
-
- Spits, H., and L.L. Lanier. 2007. Natural killer or dendritic: what's in a name? Immunity. 26:11–16. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
