

DNA damage-induced acetylation of lysine 3016 of ATM activates ATM kinase activity
- PMID: 17923702
- PMCID: PMC2169409
- DOI: 10.1128/MCB.01382-07
DNA damage-induced acetylation of lysine 3016 of ATM activates ATM kinase activity
Abstract
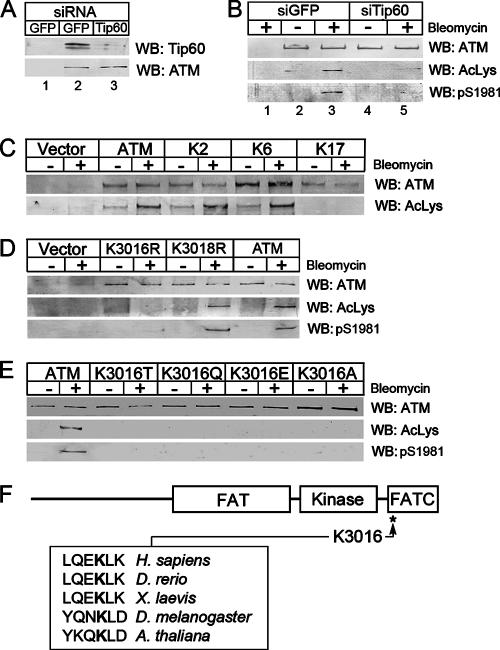
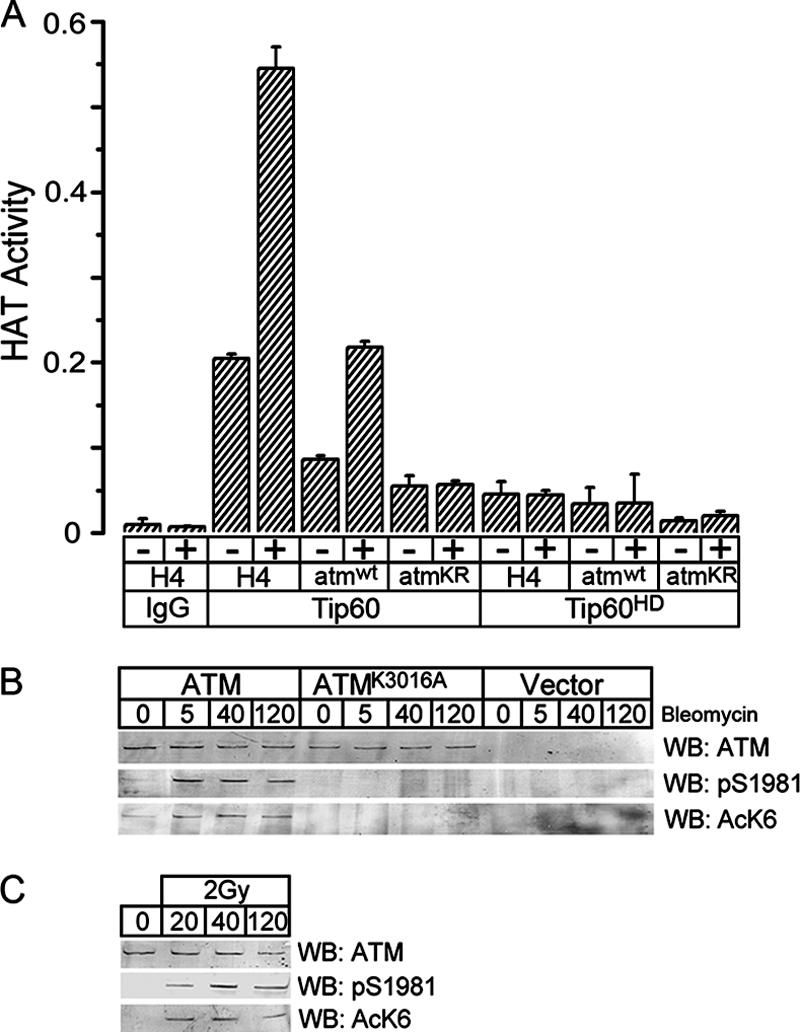
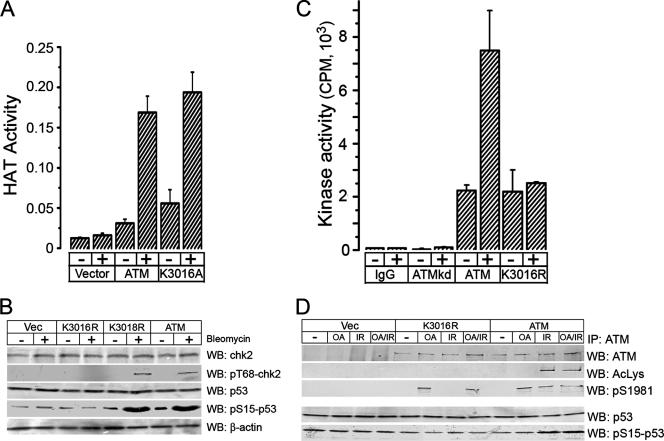
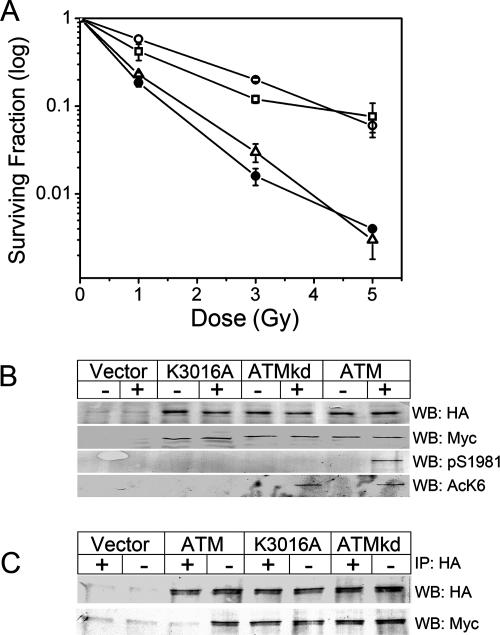
The ATM protein kinase is essential for cells to repair and survive genotoxic events. The activation of ATM's kinase activity involves acetylation of ATM by the Tip60 histone acetyltransferase. In this study, systematic mutagenesis of lysine residues was used to identify regulatory ATM acetylation sites. The results identify a single acetylation site at lysine 3016, which is located in the highly conserved C-terminal FATC domain adjacent to the kinase domain. Antibodies specific for acetyl-lysine 3016 demonstrate rapid (within 5 min) in vivo acetylation of ATM following exposure to bleomycin. Furthermore, lysine 3016 of ATM is a substrate in vitro for the Tip60 histone acetyltransferase. Mutation of lysine 3016 does not affect unstimulated ATM kinase activity but does abolish upregulation of ATM's kinase activity by DNA damage, inhibits the conversion of inactive ATM dimers to active ATM monomers, and prevents the ATM-dependent phosphorylation of the p53 and chk2 proteins. These results are consistent with a model in which acetylation of lysine 3016 in the FATC domain of ATM activates the kinase activity of ATM. The acetylation of ATM on lysine 3016 by Tip60 is therefore a key step linking the detection of DNA damage and the activation of ATM kinase activity.
Figures
References
-
- Abraham, R. T. 2004. PI 3-kinase related kinases: ‘big’ players in stress-induced signaling pathways. DNA Repair (Amsterdam) 3:883-887. - PubMed
-
- Bakkenist, C. J., and M. B. Kastan. 2003. DNA damage activates ATM through intermolecular autophosphorylation and dimer dissociation. Nature 421:499-506. - PubMed
-
- Berkovich, E., R. J. Monnat, Jr., and M. B. Kastan. 2007. Roles of ATM and NBS1 in chromatin structure modulation and DNA double-strand break repair. Nat. Cell Biol. 9:683-690. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous