

Dimensional reduction in sensorimotor systems: a framework for understanding muscle coordination of posture
- PMID: 17925254
- PMCID: PMC4121431
- DOI: 10.1016/S0079-6123(06)65019-X
Dimensional reduction in sensorimotor systems: a framework for understanding muscle coordination of posture
Abstract
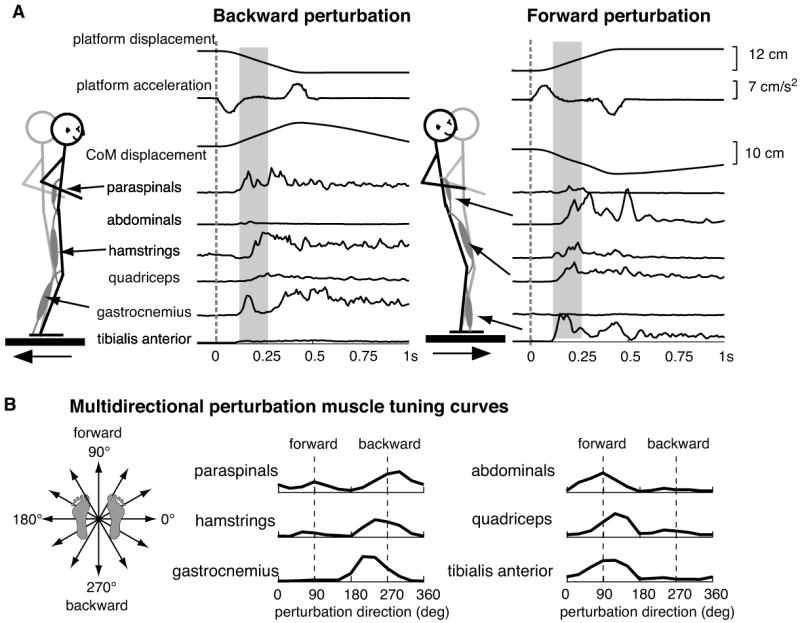
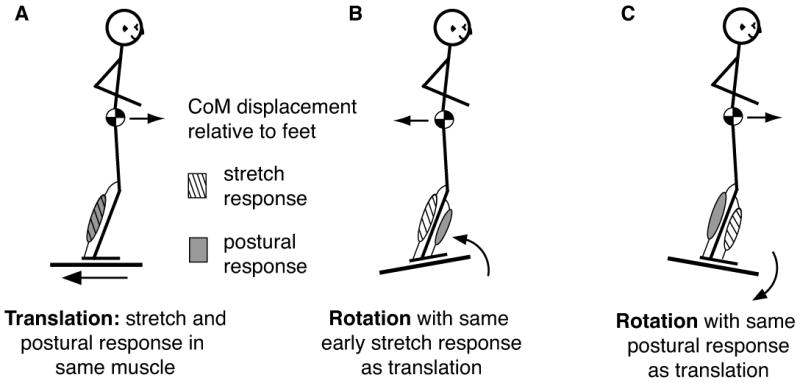
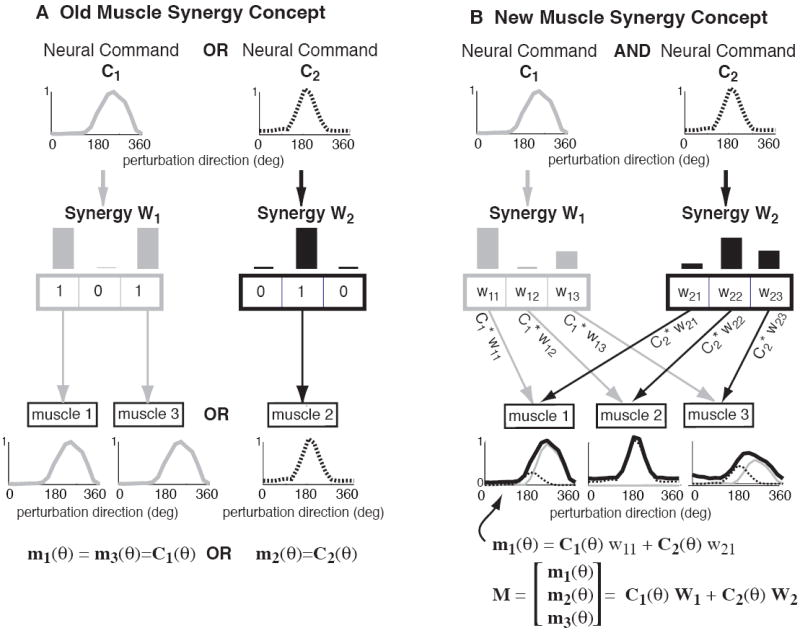
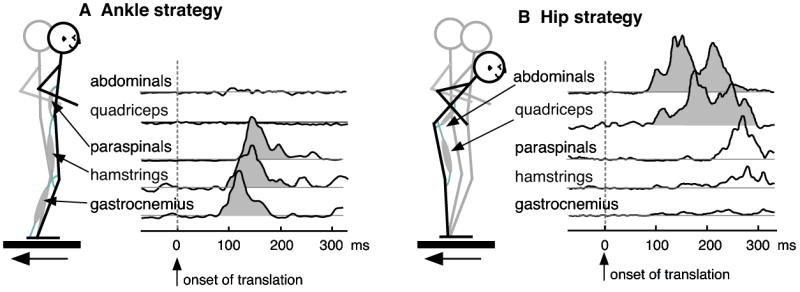
The simple act of standing up is an important and essential motor behavior that most humans and animals achieve with ease. Yet, maintaining standing balance involves complex sensorimotor transformations that must continually integrate a large array of sensory inputs and coordinate multiple motor outputs to muscles throughout the body. Multiple, redundant local sensory signals are integrated to form an estimate of a few global, task-level variables important to postural control, such as body center of mass (CoM) position and body orientation with respect to Earth-vertical. Evidence suggests that a limited set of muscle synergies, reflecting preferential sets of muscle activation patterns, are used to move task-variables such as CoM position in a predictable direction following postural perturbations. We propose a hierarchical feedback control system that allows the nervous system the simplicity of performing goal-directed computations in task-variable space, while maintaining the robustness afforded by redundant sensory and motor systems. We predict that modulation of postural actions occurs in task-variable space, and in the associated transformations between the low-dimensional task-space and high-dimensional sensor and muscle spaces. Development of neuromechanical models that reflect these neural transformations between low- and high-dimensional representations will reveal the organizational principles and constraints underlying sensorimotor transformations for balance control, and perhaps motor tasks in general. This framework and accompanying computational models could be used to formulate specific hypotheses about how specific sensory inputs and motor outputs are generated and altered following neural injury, sensory loss, or rehabilitation.
Figures
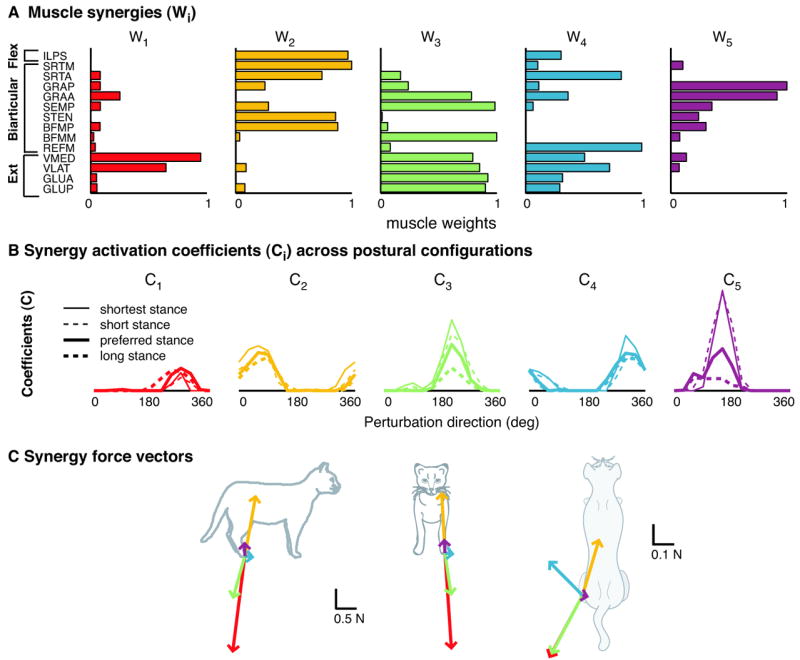
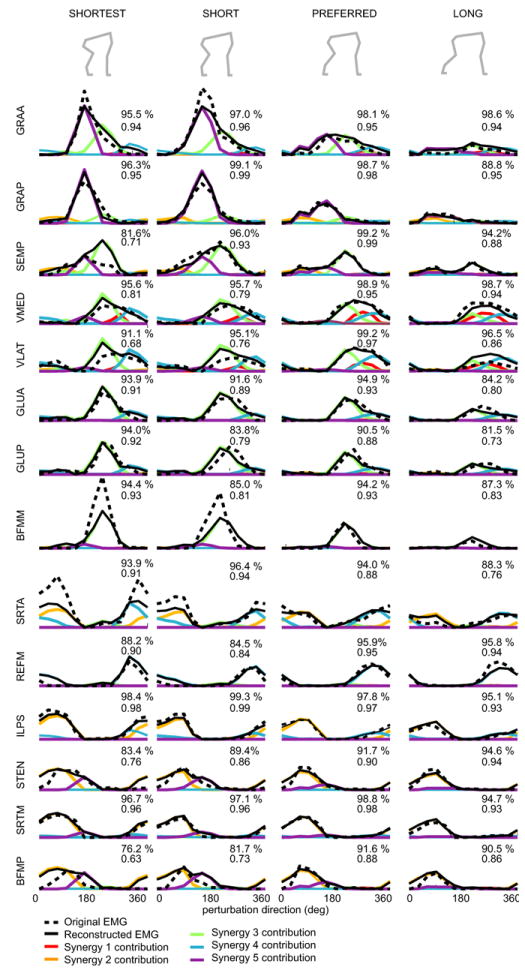
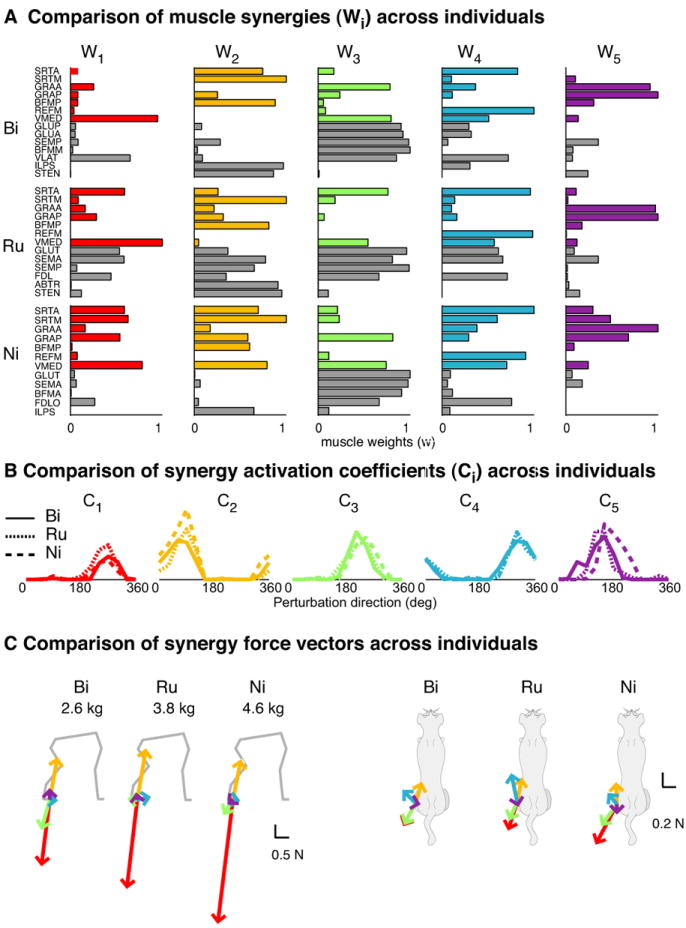
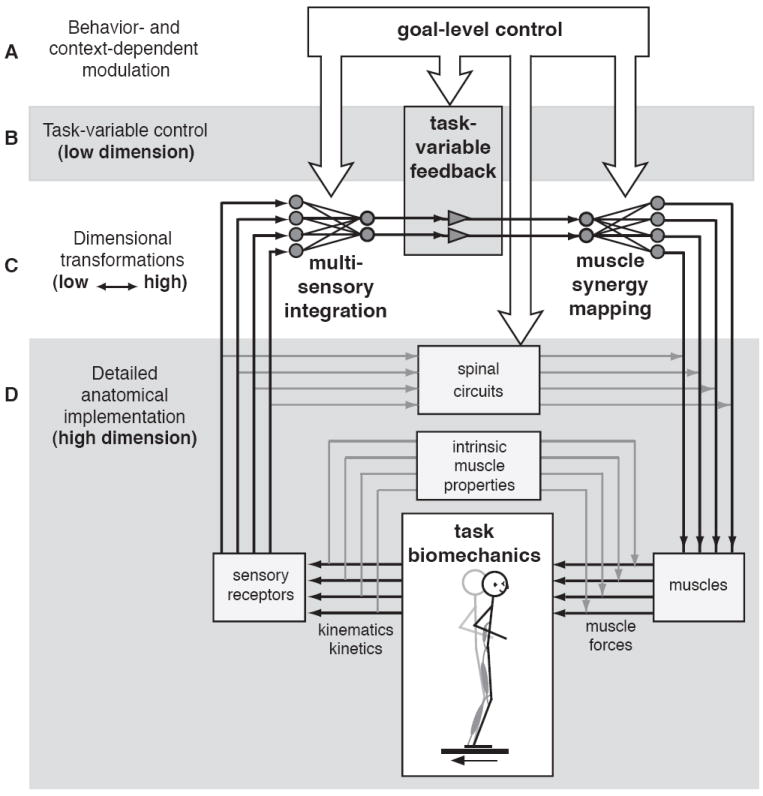
References
-
- Alexandrov AV, Frolov AA, Massion J. Biomechanical analysis of movement strategies in human forward trunk bending. I. Modeling. Biol Cybern. 2001a;84:425–34. - PubMed
-
- Alexandrov AV, Frolov AA, Massion J. Biomechanical analysis of movement strategies in human forward trunk bending. II. Experimental study. Biol Cybern. 2001b;84:435–43. - PubMed
-
- Allum JH, Bloem BR, Carpenter MG, Hulliger M, Hadders-Algra M. Proprioceptive control of posture: a review of new concepts. Gait Posture. 1998;8:214–242. - PubMed
-
- Allum JH, Carpenter MG. A speedy solution for balance and gait analysis: angular velocity measured at the centre of body mass. Curr Opin Neurol. 2005;18:15–21. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
