

Wnt and TGF-beta expression in the sponge Amphimedon queenslandica and the origin of metazoan embryonic patterning
- PMID: 17925879
- PMCID: PMC2000352
- DOI: 10.1371/journal.pone.0001031
Wnt and TGF-beta expression in the sponge Amphimedon queenslandica and the origin of metazoan embryonic patterning
Abstract
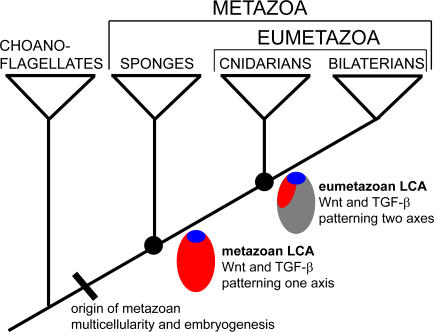
Background: The origin of metazoan development and differentiation was contingent upon the evolution of cell adhesion, communication and cooperation mechanisms. While components of many of the major cell signalling pathways have been identified in a range of sponges (phylum Porifera), their roles in development have not been investigated and remain largely unknown. Here, we take the first steps toward reconstructing the developmental signalling systems used in the last common ancestor to living sponges and eumetazoans by studying the expression of genes encoding Wnt and TGF-beta signalling ligands during the embryonic development of a sponge.
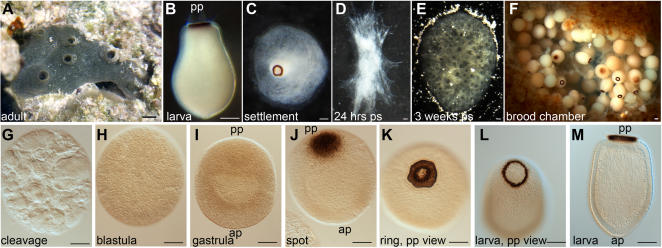
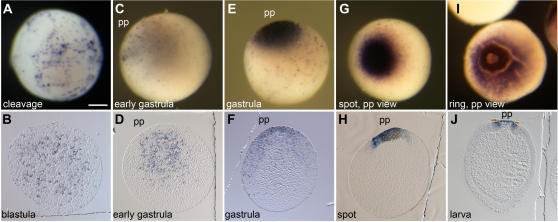
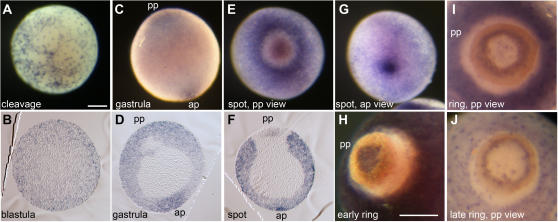
Methodology/principal findings: Using resources generated in the recent sponge Amphimedon queenslandica (Demospongiae) genome project, we have recovered genes encoding Wnt and TGF-beta signalling ligands that are critical in patterning metazoan embryos. Both genes are expressed from the earliest stages of Amphimedon embryonic development in highly dynamic patterns. At the time when the Amphimedon embryos begin to display anterior-posterior polarity, Wnt expression becomes localised to the posterior pole and this expression continues until the swimming larva stage. In contrast, TGF-beta expression is highest at the anterior pole. As in complex animals, sponge Wnt and TGF-beta expression patterns intersect later in development during the patterning of a sub-community of cells that form a simple tissue-like structure, the pigment ring. Throughout development, Wnt and TGF-beta are expressed radially along the anterior-posterior axis.
Conclusions/significance: We infer from the expression of Wnt and TGF-beta in Amphimedon that the ancestor that gave rise to sponges, cnidarians and bilaterians had already evolved the capacity to direct the formation of relatively sophisticated body plans, with axes and tissues. The radially symmetrical expression patterns of Wnt and TGF-beta along the anterior-posterior axis of sponge embryos and larvae suggest that these signalling pathways contributed to establishing axial polarity in the very first metazoans.
Conflict of interest statement
Figures
Similar articles
-
Structure and expression of conserved Wnt pathway components in the demosponge Amphimedon queenslandica.Evol Dev. 2010 Sep-Oct;12(5):494-518. doi: 10.1111/j.1525-142X.2010.00435.x. Evol Dev. 2010. PMID: 20883218
-
Surprisingly rich repertoire of Wnt genes in the demosponge Halisarca dujardini.BMC Evol Biol. 2016 Jun 10;16(1):123. doi: 10.1186/s12862-016-0700-6. BMC Evol Biol. 2016. PMID: 27287511 Free PMC article.
-
WNT/beta-catenin signalling and epithelial patterning in the homoscleromorph sponge Oscarella.PLoS One. 2009 Jun 8;4(6):e5823. doi: 10.1371/journal.pone.0005823. PLoS One. 2009. PMID: 19503791 Free PMC article.
-
From traveler to homebody: Which signaling mechanisms sponge larvae use to become adult sponges?Adv Protein Chem Struct Biol. 2019;116:421-449. doi: 10.1016/bs.apcsb.2019.02.002. Epub 2019 Mar 14. Adv Protein Chem Struct Biol. 2019. PMID: 31036299 Review.
-
Heads or tails? Amphioxus and the evolution of anterior-posterior patterning in deuterostomes.Dev Biol. 2002 Jan 15;241(2):209-28. doi: 10.1006/dbio.2001.0503. Dev Biol. 2002. PMID: 11784106 Review.
Cited by
-
Involvement of Wnt signaling pathways in the metamorphosis of the bryozoan Bugula neritina.PLoS One. 2012;7(3):e33323. doi: 10.1371/journal.pone.0033323. Epub 2012 Mar 20. PLoS One. 2012. PMID: 22448242 Free PMC article.
-
Origin and evolution of the Notch signalling pathway: an overview from eukaryotic genomes.BMC Evol Biol. 2009 Oct 13;9:249. doi: 10.1186/1471-2148-9-249. BMC Evol Biol. 2009. PMID: 19825158 Free PMC article.
-
Exploring genome gene content and morphological analysis to test recalcitrant nodes in the animal phylogeny.PLoS One. 2023 Mar 23;18(3):e0282444. doi: 10.1371/journal.pone.0282444. eCollection 2023. PLoS One. 2023. PMID: 36952565 Free PMC article.
-
The transcription factor NF-kappaB in the demosponge Amphimedon queenslandica: insights on the evolutionary origin of the Rel homology domain.Dev Genes Evol. 2008 Jan;218(1):23-32. doi: 10.1007/s00427-007-0197-5. Epub 2008 Jan 4. Dev Genes Evol. 2008. PMID: 18175145
-
Gene classification based on amino acid motifs and residues: the DLX (distal-less) test case.PLoS One. 2009 Jun 1;4(6):e5748. doi: 10.1371/journal.pone.0005748. PLoS One. 2009. PMID: 19484130 Free PMC article.
References
-
- Martindale MQ. The evolution of metazoan axial properties. Nat Rev Genet. 2005;6:917–27. - PubMed
-
- Erwin DH, Davidson EH. The last common bilaterian ancestor. Development. 2002;129:3021–32. - PubMed
-
- Lee PN, Pang K, Matus DQ, Martindale MQ. A WNT of things to come: evolution of Wnt signaling and polarity in cnidarians. Semin Cell Dev Biol. 2006;17:157–67. - PubMed
-
- Miller DJ, Ball EE, Technau U. Cnidarians and ancestral genetic complexity in the animal kingdom. Trends Genet. 2005;21:536–9. - PubMed
-
- Kusserow A, Pang K, Sturm C, Hrouda M, Lentfer J, et al. Unexpected complexity of the Wnt gene family in a sea anemone. Nature. 2005;433:156–60. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
