

SHP-2 is required for the maintenance of cardiac progenitors
- PMID: 17928416
- PMCID: PMC2807747
- DOI: 10.1242/dev.009290
SHP-2 is required for the maintenance of cardiac progenitors
Abstract
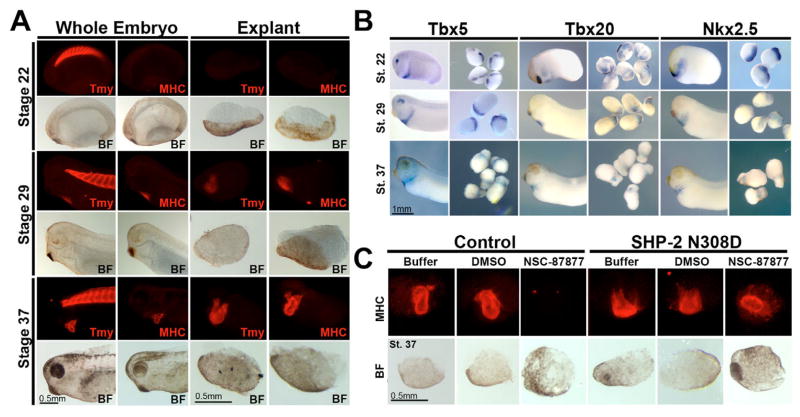
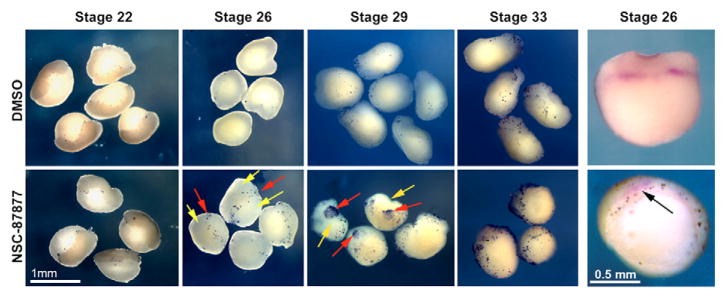
The isolation and culturing of cardiac progenitor cells has demonstrated that growth factor signaling is required to maintain cardiac cell survival and proliferation. In this study, we demonstrate in Xenopus that SHP-2 activity is required for the maintenance of cardiac precursors in vivo. In the absence of SHP-2 signaling, cardiac progenitor cells downregulate genes associated with early heart development and fail to initiate cardiac differentiation. We further show that this requirement for SHP-2 is restricted to cardiac precursor cells undergoing active proliferation. By demonstrating that SHP-2 is phosphorylated on Y542/Y580 and that it binds to FRS-2, we place SHP-2 in the FGF pathway during early embryonic heart development. Furthermore, we demonstrate that inhibition of FGF signaling mimics the cellular and biochemical effects of SHP-2 inhibition and that these effects can be rescued by constitutively active/Noonan-syndrome-associated forms of SHP-2. Collectively, these results show that SHP-2 functions within the FGF/MAPK pathway to maintain survival of proliferating populations of cardiac progenitor cells.
Figures






Similar articles
-
SHP-2 acts via ROCK to regulate the cardiac actin cytoskeleton.Development. 2012 Mar;139(5):948-57. doi: 10.1242/dev.067579. Epub 2012 Jan 25. Development. 2012. PMID: 22278918 Free PMC article.
-
Frs2alpha-deficiency in cardiac progenitors disrupts a subset of FGF signals required for outflow tract morphogenesis.Development. 2008 Nov;135(21):3611-22. doi: 10.1242/dev.025361. Epub 2008 Oct 2. Development. 2008. PMID: 18832393 Free PMC article.
-
FRS2α-mediated FGF signals suppress premature differentiation of cardiac stem cells through regulating autophagy activity.Circ Res. 2012 Feb 17;110(4):e29-39. doi: 10.1161/CIRCRESAHA.111.255950. Epub 2011 Dec 29. Circ Res. 2012. PMID: 22207710 Free PMC article.
-
SHP-2 regulates growth factor dependent vascular signalling and function.Mini Rev Med Chem. 2014;14(6):471-83. doi: 10.2174/1389557514999140506094738. Mini Rev Med Chem. 2014. PMID: 22512561 Review.
-
Signaling pathways controlling second heart field development.Circ Res. 2009 Apr 24;104(8):933-42. doi: 10.1161/CIRCRESAHA.109.194464. Circ Res. 2009. PMID: 19390062 Review.
Cited by
-
Vascular mechanotransduction data in a rodent model of diabetes: Pressure-induced regulation of SHP2 and associated signaling in the rat inferior vena cava.Data Brief. 2017 Sep 20;15:300-307. doi: 10.1016/j.dib.2017.09.028. eCollection 2017 Dec. Data Brief. 2017. PMID: 29214191 Free PMC article.
-
Expanding the genetic toolkit in Xenopus: Approaches and opportunities for human disease modeling.Dev Biol. 2017 Jun 15;426(2):325-335. doi: 10.1016/j.ydbio.2016.04.009. Epub 2016 Apr 22. Dev Biol. 2017. PMID: 27109192 Free PMC article. Review.
-
Proteomic Analysis of Myocardia Containing the Obscurin R4344Q Mutation Linked to Hypertrophic Cardiomyopathy.Front Physiol. 2020 May 18;11:478. doi: 10.3389/fphys.2020.00478. eCollection 2020. Front Physiol. 2020. PMID: 32528308 Free PMC article.
-
DDIT3 is associated with breast cancer prognosis and immune microenvironment: an integrative bioinformatic and immunohistochemical analysis.J Cancer. 2024 May 28;15(12):3873-3889. doi: 10.7150/jca.96491. eCollection 2024. J Cancer. 2024. PMID: 38911383 Free PMC article.
-
Focal adhesion kinase is essential for cardiac looping and multichamber heart formation.Genesis. 2010 Aug;48(8):492-504. doi: 10.1002/dvg.20650. Genesis. 2010. PMID: 20572259 Free PMC article.
References
-
- Alexander J, Rothenberg M, Henry GL, Stainier DY. casanova plays an early and essential role in endoderm formation in zebrafish. Dev Biol. 1999;215:343–357. - PubMed
-
- Alexiades MR, Cepko C. Quantitative analysis of proliferation and cell cycle length during development of the rat retina. Dev Dyn. 1996;205:293–307. - PubMed
-
- Allanson J. The first Noonan syndrome gene: PTPN11, which encodes the protein tyrosine phosphatase SHP-2. Pediatr Res. 2002;52:471. - PubMed
-
- Alsan BH, Schultheiss TM. Regulation of avian cardiogenesis by Fgf8 signaling. Development. 2002;129:1935–1943. - PubMed
-
- Araki T, Mohi MG, Ismat FA, Bronson RT, Williams IR, Kutok JL, Yang W, Pao LI, Gilliland DG, Epstein JA, et al. Mouse model of Noonan syndrome reveals cell type- and gene dosage-dependent effects of Ptpn11 mutation. Nat Med. 2004;10:849–857. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
