

Modular control of limb movements during human locomotion
- PMID: 17928457
- PMCID: PMC6672838
- DOI: 10.1523/JNEUROSCI.2644-07.2007
Modular control of limb movements during human locomotion
Abstract
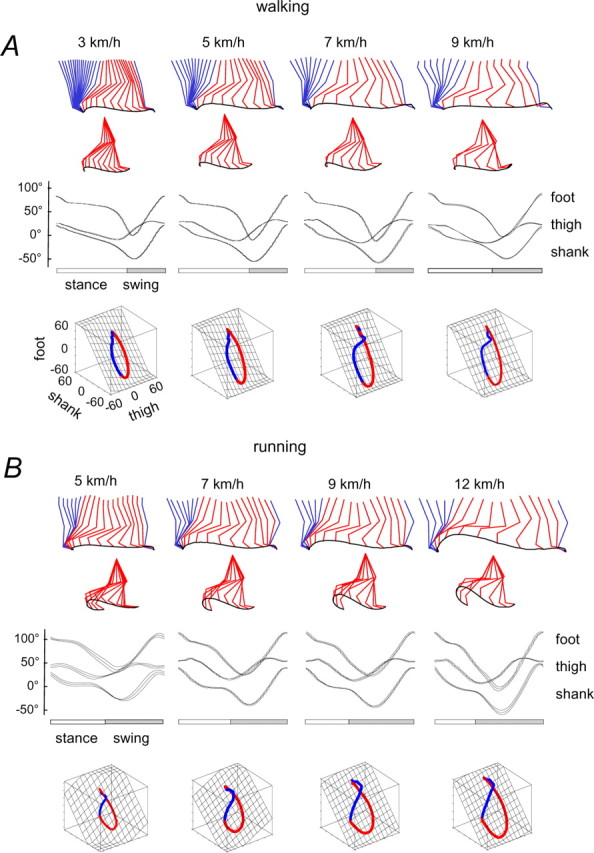
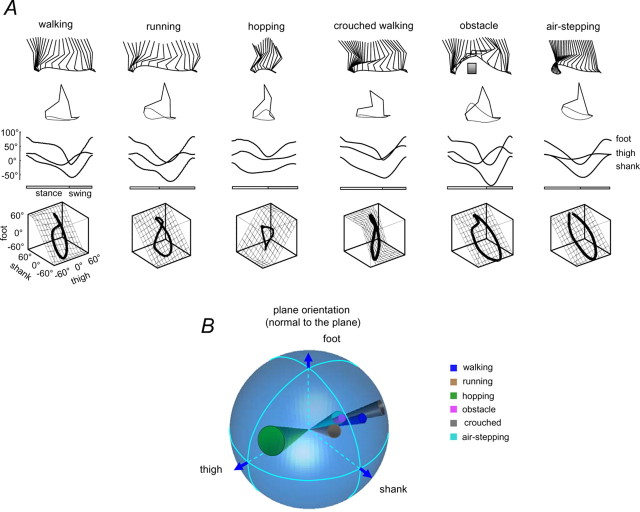
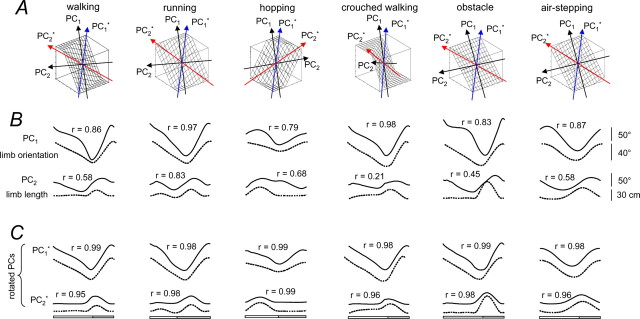
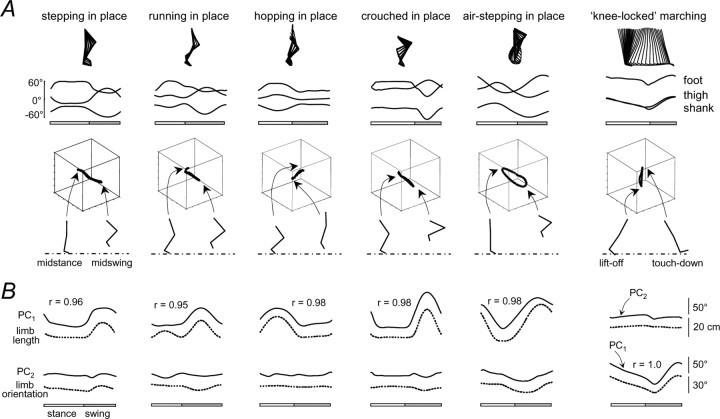
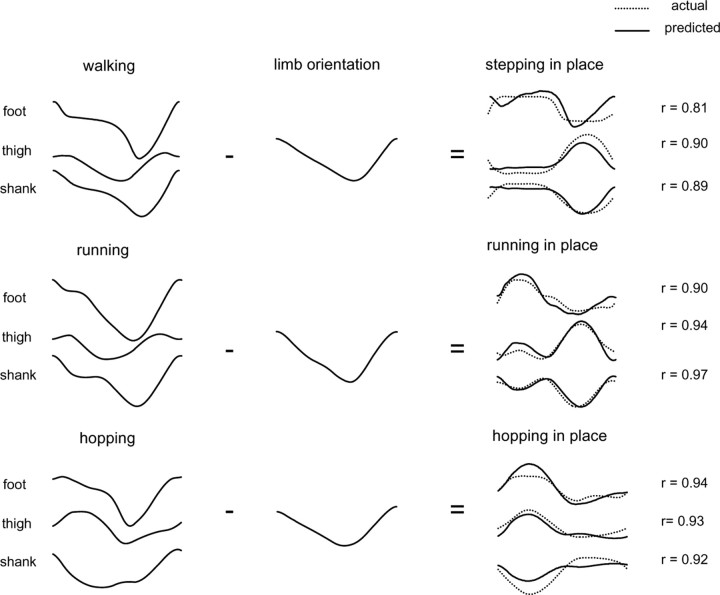
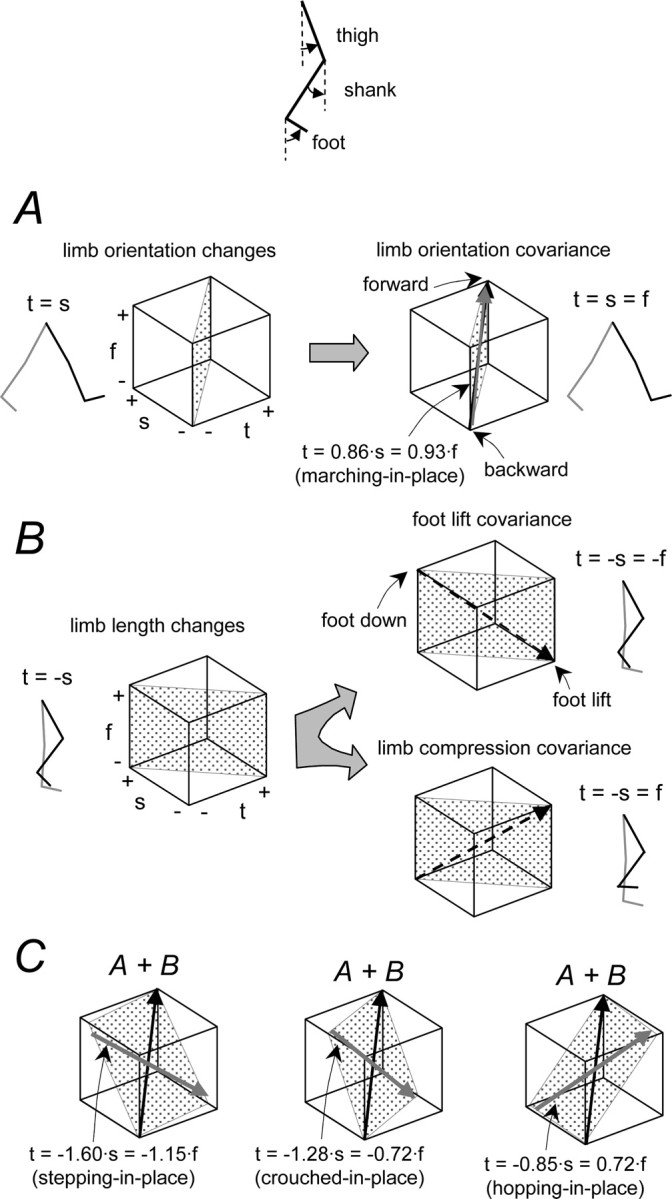
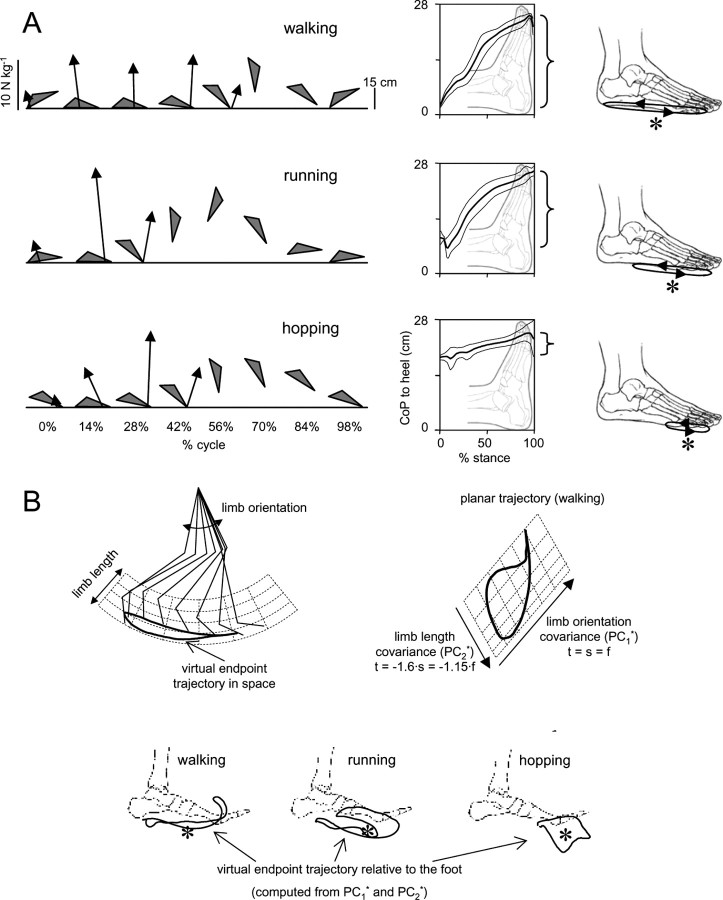
The idea that the CNS may control complex interactions by modular decomposition has received considerable attention. We explored this idea for human locomotion by examining limb kinematics. The coordination of limb segments during human locomotion has been shown to follow a planar law for walking at different speeds, directions, and levels of body unloading. We compared the coordination for different gaits. Eight subjects were asked to walk and run on a treadmill at different speeds or to walk, run, and hop over ground at a preferred speed. To explore various constraints on limb movements, we also recorded stepping over an obstacle, walking with the knees flexed, and air-stepping with body weight support. We found little difference among covariance planes that depended on speed, but there were differences that depended on gait. In each case, we could fit the planar trajectories with a weighted sum of the limb length and orientation trajectories. This suggested that limb length and orientation might provide independent predictors of limb coordination. We tested this further by having the subjects step, run, and hop in place, thereby varying only limb length and maintaining limb orientation fixed, and also by marching with knees locked to maintain limb length constant while varying orientation. The results were consistent with a modular control of limb kinematics where limb movements result from a superposition of separate length- and orientation-related angular covariance. The hypothesis finds support in the animal findings that limb proprioception may also be encoded in terms of these global limb parameters.
Figures
Similar articles
-
On the origin of planar covariation of elevation angles during human locomotion.J Neurophysiol. 2008 Apr;99(4):1890-8. doi: 10.1152/jn.01308.2007. Epub 2008 Feb 13. J Neurophysiol. 2008. PMID: 18272871
-
Walk-, run- and gallop-like gait patterns in human sideways locomotion.J Electromyogr Kinesiol. 2013 Dec;23(6):1480-4. doi: 10.1016/j.jelekin.2013.08.005. Epub 2013 Aug 30. J Electromyogr Kinesiol. 2013. PMID: 24055531
-
Cerebellar damage produces context-dependent deficits in control of leg dynamics during obstacle avoidance.Exp Brain Res. 2004 May;156(2):149-63. doi: 10.1007/s00221-003-1776-y. Epub 2004 Jan 31. Exp Brain Res. 2004. PMID: 14758452
-
Kinematic control of walking.Arch Ital Biol. 2002 Oct;140(4):263-72. Arch Ital Biol. 2002. PMID: 12228979 Review.
-
Contributions to the understanding of gait control.Dan Med J. 2014 Apr;61(4):B4823. Dan Med J. 2014. PMID: 24814597 Review.
Cited by
-
Muscle synergies and spinal maps are sensitive to the asymmetry induced by a unilateral stroke.J Neuroeng Rehabil. 2015 Apr 18;12:39. doi: 10.1186/s12984-015-0031-7. J Neuroeng Rehabil. 2015. PMID: 25928264 Free PMC article.
-
Effects of visual deprivation on intra-limb coordination during walking in children and adults.Exp Brain Res. 2009 Sep;198(1):95-106. doi: 10.1007/s00221-009-1937-8. Epub 2009 Jul 18. Exp Brain Res. 2009. PMID: 19618172
-
The Effects of 12-Week Dual-Task Physical-Cognitive Training on Gait, Balance, Lower Extremity Muscle Strength, and Cognition in Older Adult Women: A Randomized Study.Int J Environ Res Public Health. 2023 Apr 13;20(8):5498. doi: 10.3390/ijerph20085498. Int J Environ Res Public Health. 2023. PMID: 37107780 Free PMC article. Clinical Trial.
-
Empirical Evaluation of Voluntarily Activatable Muscle Synergies.Front Comput Neurosci. 2017 Sep 6;11:82. doi: 10.3389/fncom.2017.00082. eCollection 2017. Front Comput Neurosci. 2017. PMID: 28932190 Free PMC article.
-
Effects of Hemispheric Stroke Localization on the Reorganization of Arm Movements within Different Mechanical Environments.Life (Basel). 2021 Apr 23;11(5):383. doi: 10.3390/life11050383. Life (Basel). 2021. PMID: 33922668 Free PMC article.
References
-
- Arampatzis A, Bruggemann GP, Metzler V. The effect of speed on leg stiffness and joint kinetics in human running. J Biomech. 1999;32:1349–1353. - PubMed
-
- Batschelet E. New York: Academic; 1981. Circular statistics in biology.
-
- Bernstein N. Oxford: Pergamon; 1967. The co-ordination and regulation of movements.
-
- Bianchi L, Angelini D, Lacquaniti F. Individual characteristics of human walking mechanics. Pflügers Arch. 1998a;436:343–356. - PubMed
-
- Bianchi L, Angelini D, Orani GP, Lacquaniti F. Kinematic co-ordination in human gait: relation to mechanical energy cost. J Neurophysiol. 1998b;79:2155–2170. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources