

Characterization of a natural heterodimer between MLV genomic RNA and the SD' retroelement generated by alternative splicing
- PMID: 17928575
- PMCID: PMC2080594
- DOI: 10.1261/rna.713807
Characterization of a natural heterodimer between MLV genomic RNA and the SD' retroelement generated by alternative splicing
Abstract
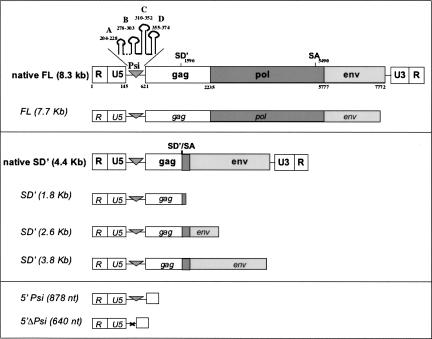
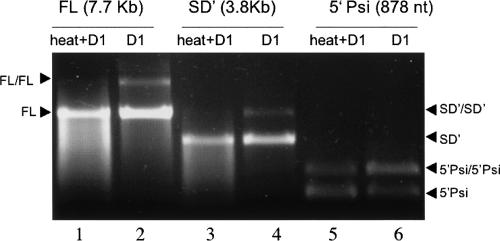
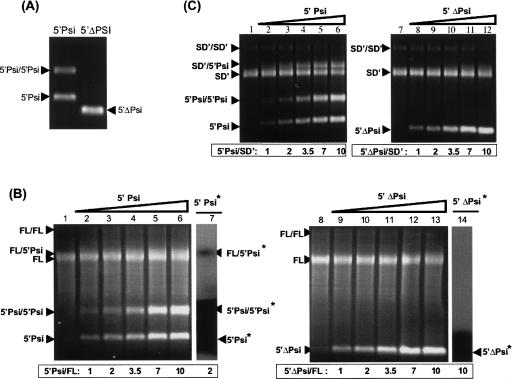
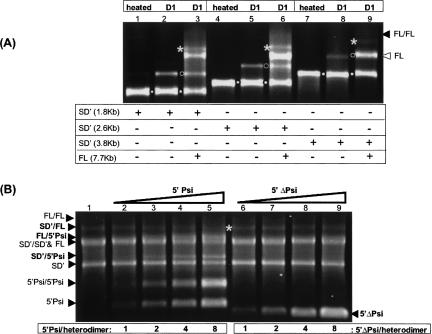
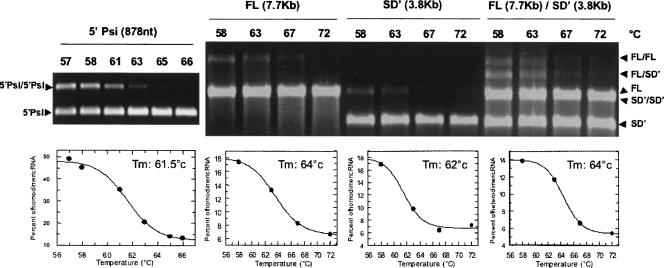
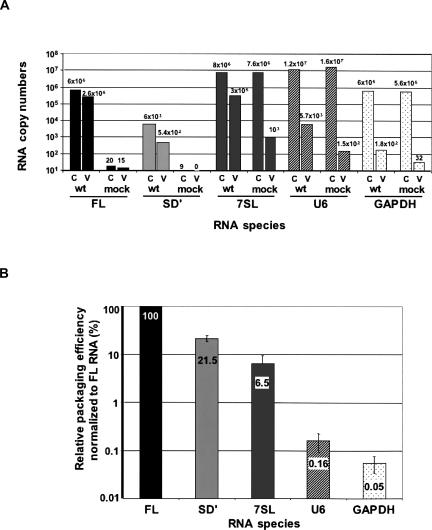
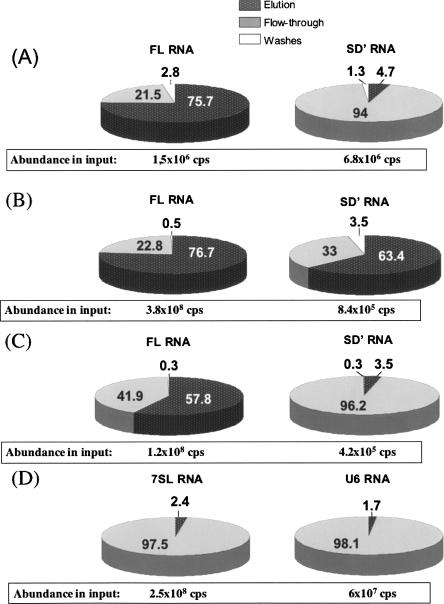
Murine leukemia virus (MLV) specifically packages both genomic RNA (FL RNA) and a subgenomic RNA, which we call SD'. SD' RNA results from alternative splicing of FL RNA. It is reverse-transcribed, and its DNA copy, integrated into the host genome, constitutes a splice donor-associated retroelement. FL and SD' RNAs share a common 5'-UTR that includes the packaging/dimerization signal (Psi). To investigate whether the mechanism of copackaging of these two RNAs involves RNA heterodimerization, we examined the spontaneous dimerization capacity of the two RNAs as large synthetic RNAs transcribed in vitro. We showed that SD' RNA not only formed homodimers with similar efficiency as the FL RNA, but that FL and SD' RNAs also formed FL/SD' heterodimers via Psi sequences. Comparison of the thermostabilities determined for these different dimeric species and competition experiments with Psi RNA fragments indicate the recruitment of similar dimer-linkage interactions within the Psi region. To validate these results, the dimeric state of the SD' RNA was analyzed in MLV particles. RNA capture assays performed with the FL RNA as bait revealed that SD', and not the host packageable U6 or 7SL RNAs, was associated with the FL RNA in virions. Heterodimerization of SD' RNA with FL RNA may argue for the recent concept of a nuclear dimerization at or near the site of transcription and raises the new hypothesis of RNA dimerization during splicing. Furthermore, FL/SD' heterodimerization may have leukemogenic consequences by influencing the pool of genomic dimers that will undergo recombinogenic template switching by reverse transcriptase.
Figures
Similar articles
-
Murine leukemia virus RNA dimerization is coupled to transcription and splicing processes.Retrovirology. 2010 Aug 5;7:64. doi: 10.1186/1742-4690-7-64. Retrovirology. 2010. PMID: 20687923 Free PMC article.
-
A new retroelement constituted by a natural alternatively spliced RNA of murine replication-competent retroviruses.EMBO J. 2003 Sep 15;22(18):4866-75. doi: 10.1093/emboj/cdg450. EMBO J. 2003. PMID: 12970198 Free PMC article.
-
Nonrandom dimerization of murine leukemia virus genomic RNAs.J Virol. 2004 Nov;78(22):12129-39. doi: 10.1128/JVI.78.22.12129-12139.2004. J Virol. 2004. PMID: 15507599 Free PMC article.
-
Insights into the nuclear export of murine leukemia virus intron-containing RNA.RNA Biol. 2015;12(9):942-9. doi: 10.1080/15476286.2015.1065375. RNA Biol. 2015. PMID: 26158194 Free PMC article. Review.
-
[Analysis on primate lentivirus genome dimerization in virion].Uirusu. 2005 Jun;55(1):153-60. doi: 10.2222/jsv.55.153. Uirusu. 2005. PMID: 16308542 Review. Japanese.
Cited by
-
Cell biology of retroviral RNA packaging.RNA Biol. 2011 Jul-Aug;8(4):572-80. doi: 10.4161/rna.8.4.16030. Epub 2011 Jul 1. RNA Biol. 2011. PMID: 21691151 Free PMC article. Review.
-
Murine leukemia virus (MLV) P50 protein induces cell transformation via transcriptional regulatory function.Retrovirology. 2023 Sep 12;20(1):16. doi: 10.1186/s12977-023-00631-w. Retrovirology. 2023. PMID: 37700325 Free PMC article.
-
NXF1 and CRM1 nuclear export pathways orchestrate nuclear export, translation and packaging of murine leukaemia retrovirus unspliced RNA.RNA Biol. 2020 Apr;17(4):528-538. doi: 10.1080/15476286.2020.1713539. Epub 2020 Jan 23. RNA Biol. 2020. PMID: 31918596 Free PMC article.
-
Intact Viral Particle Counts Measured by Flow Virometry Provide Insight into the Infectivity and Genome Packaging Efficiency of Moloney Murine Leukemia Virus.J Virol. 2020 Jan 6;94(2):e01600-19. doi: 10.1128/JVI.01600-19. Print 2020 Jan 6. J Virol. 2020. PMID: 31694951 Free PMC article.
-
Visualization of Retroviral Gag-Genomic RNA Cellular Interactions Leading to Genome Encapsidation and Viral Assembly: An Overview.Viruses. 2022 Feb 5;14(2):324. doi: 10.3390/v14020324. Viruses. 2022. PMID: 35215917 Free PMC article. Review.
References
-
- Aagaard, L., Rasmussen, S.V., Mikkelsen, J.G., Pedersen, F.S. Efficient replication of full-length murine leukemia viruses modified at the dimer initiation site regions. Virology. 2004;318:360–370. - PubMed
-
- Bonnet-Mathoniere, B., Girard, P.M., Muriaux, D., Paoletti, J. Nucleocapsid protein 10 activates dimerization of the RNA of Moloney murine leukaemia virus in vitro. Eur. J. Biochem. 1996;238:129–135. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous