

CBP Is a dosage-dependent regulator of nuclear factor-kappaB suppression by the estrogen receptor
- PMID: 17932106
- PMCID: PMC2234588
- DOI: 10.1210/me.2007-0324
CBP Is a dosage-dependent regulator of nuclear factor-kappaB suppression by the estrogen receptor
Abstract
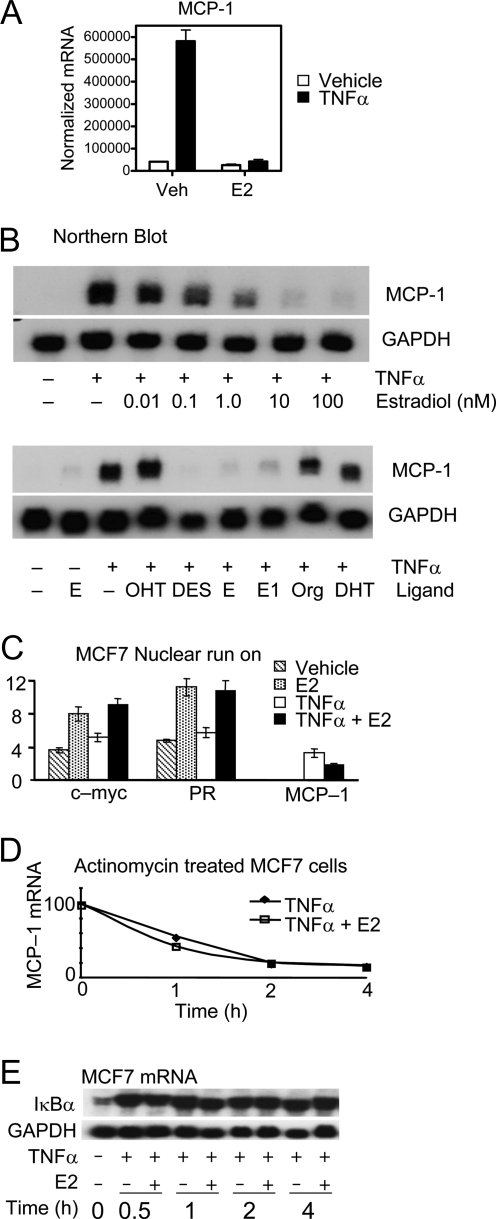
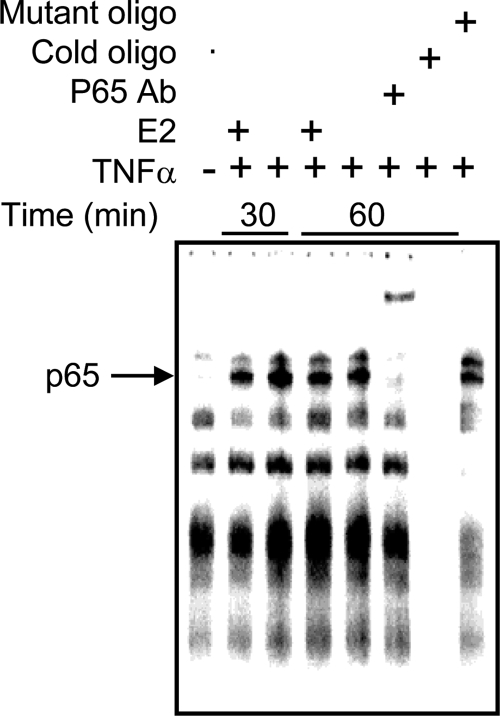
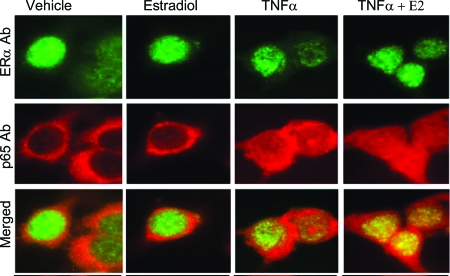
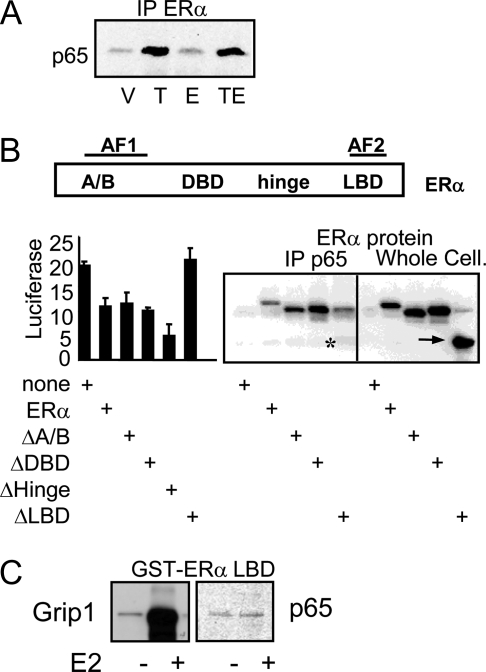
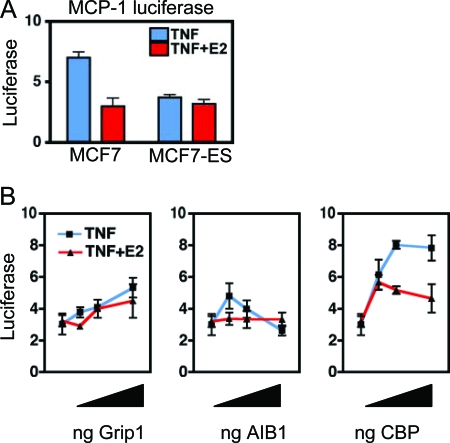
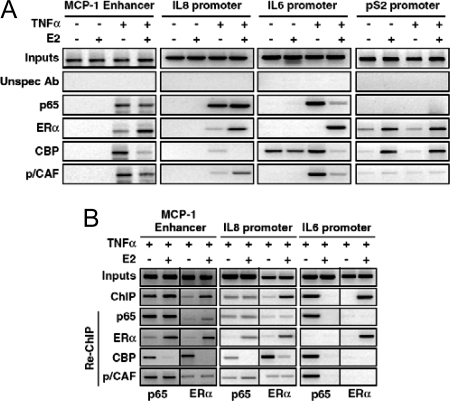
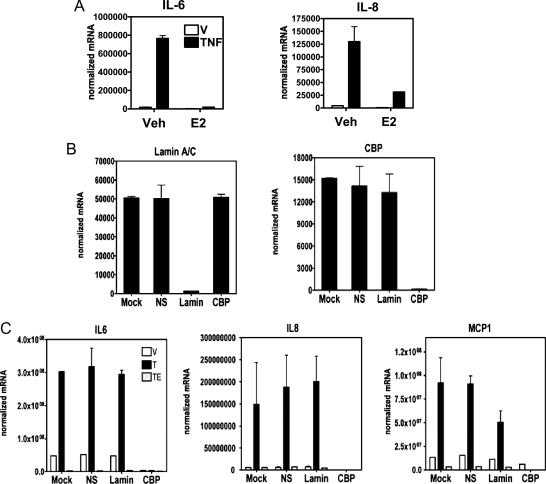
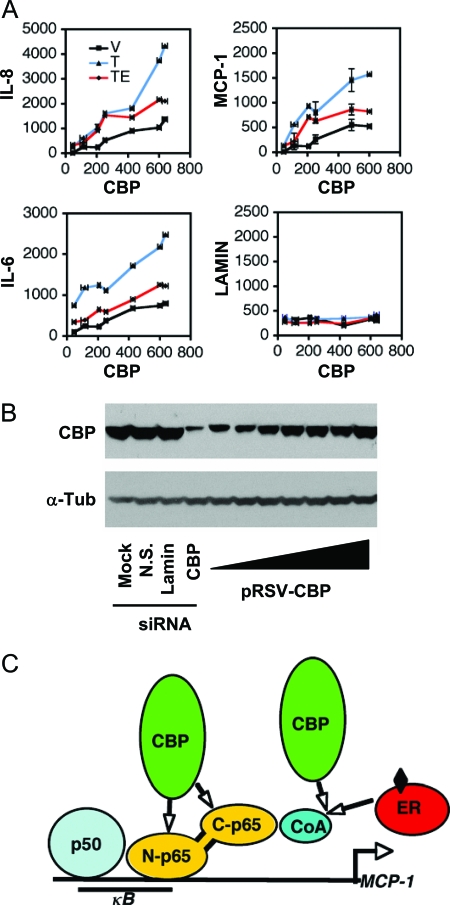
The estrogen receptor (ER) protects against debilitating effects of the inflammatory response by inhibiting the proinflammatory transcription factor nuclear factor-kappaB (NFkappaB). Heretofore cAMP response element-binding protein (CREB)-binding protein (CBP) has been suggested to mediate inhibitory cross talk by functioning either as a scaffold that links ER and NFkappaB or as a required cofactor that competitively binds to one or the other transcriptional factor. However, here we demonstrate that ER is recruited to the NFkappaB response element of the MCP-1 (monocyte chemoattractant protein-1) and IL-8 promoters and displaces CBP, but not p65, in the MCF-7 breast cancer cell line. In contrast, ER displaced p65 and associated coregulators from the IL-6 promoter, demonstrating a gene-specific role for CBP in integrating inflammatory and steroid signaling. Further, RNA interference and overexpression studies demonstrated that CBP dosage regulates estrogen-mediated suppression of MCP-1 and IL-8, but not IL-6, gene expression. This work further demonstrates that CBP dosage is a critical regulator of gene-specific signal integration between the ER- and NFkappaB-signaling pathways.
Figures
References
-
- Koehler KF, Helguero LA, Haldosen LA, Warner M, Gustafsson JA 2005 Reflections on the discovery and significance of estrogen receptor β. Endocr Rev 26:465–478 - PubMed
-
- Chadwick CC, Chippari S, Matelan E, Borges-Marcucci L, Eckert AM, Keith Jr JC, Albert LM, Leathurby Y, Harris HA, Bhat RA, Ashwell M, Trybulski E, Winneker RC, Adelman SJ, Steffan RJ, Harnish DC 2005 Identification of pathway-selective estrogen receptor ligands that inhibit NF-κB transcriptional activity. Proc Natl Acad Sci USA 102:2543–2548 - PMC - PubMed
-
- Jimi E, Ghosh S 2005 Role of nuclear factor-κB in the immune system and bone. Immunol Rev 208:80–87 - PubMed
-
- Ghosh S, Karin M 2002 Missing pieces in the NF-κB puzzle. Cell 109 (Suppl):S81–S96 - PubMed
-
- Hayden MS, Ghosh S 2004 Signaling to NF-κB. Genes Dev 18:2195–2224 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
