

Olfactory marker protein modulates the cAMP kinetics of the odour-induced response in cilia of mouse olfactory receptor neurons
- PMID: 17932148
- PMCID: PMC2375515
- DOI: 10.1113/jphysiol.2007.142471
Olfactory marker protein modulates the cAMP kinetics of the odour-induced response in cilia of mouse olfactory receptor neurons
Abstract
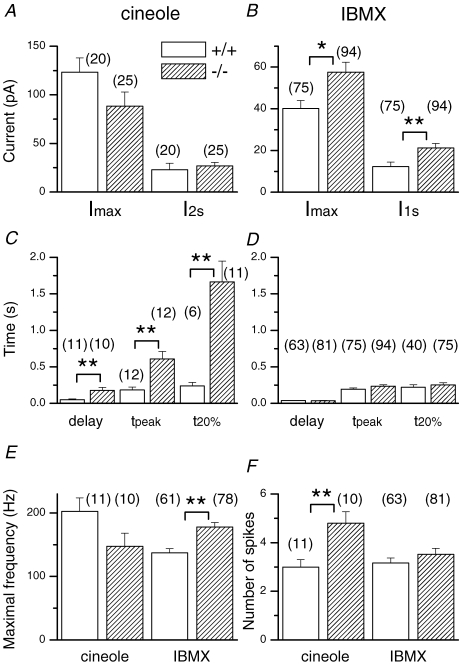
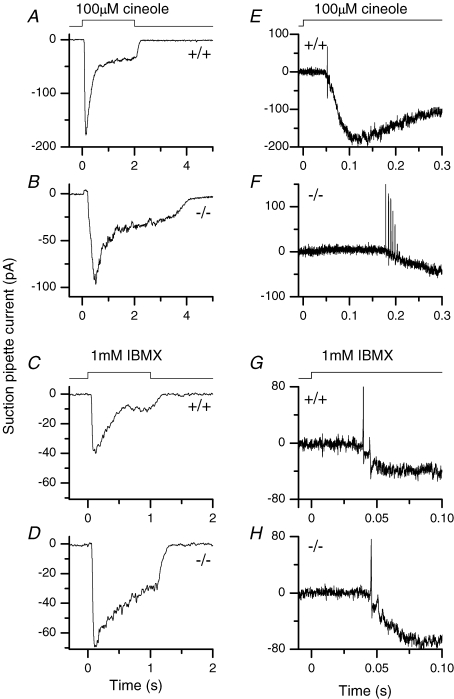
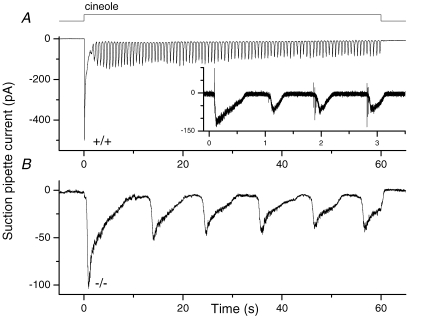
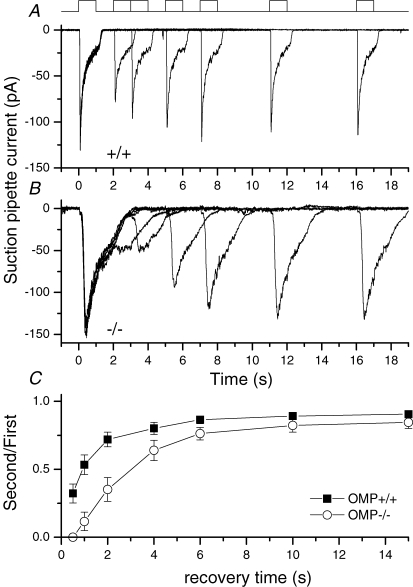
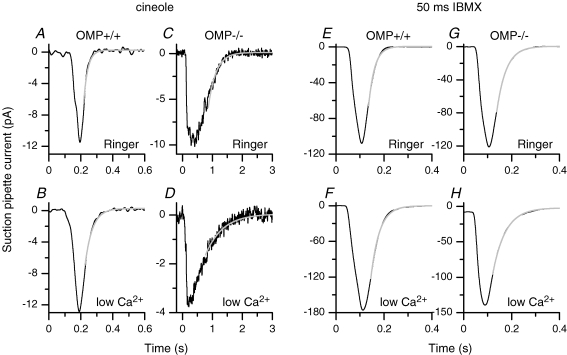
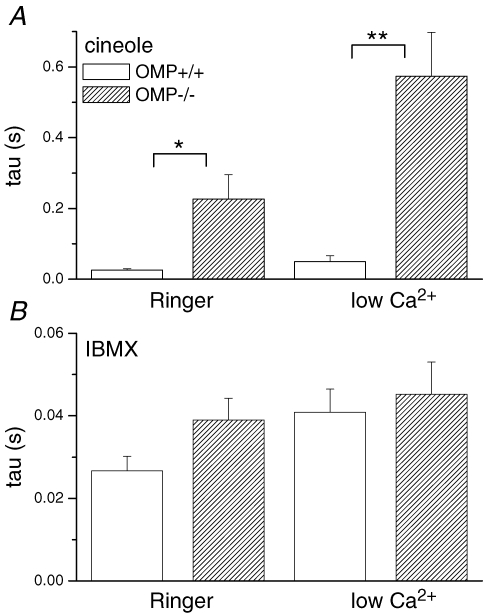
Olfactory marker protein (OMP), a phylogenetically conserved protein, is highly, and almost exclusively, expressed in vertebrate olfactory receptor neurons (ORNs). Although OMP is widely used as a marker for ORNs, its function has remained largely elusive. Here we used suction-pipette recordings from isolated ORNs of OMP(-/-) mice to investigate its role in olfactory transduction. Vertebrate olfactory transduction is initiated when odourants bind to receptor proteins to activate an adenylyl cyclase via a G protein-coupled signalling pathway. This leads to an increase in cAMP and the opening of a cyclic nucleotide-gated (CNG), non-selective cation channel which depolarizes the cells. Ca(2+) influx through the CNG channel in turn activates a Ca(2+)-activated Cl(-) channel, causing a Cl(-) efflux and further depolarization. In the absence of OMP, the time-to-transient-peak of the response, the latency to first spike, and the response termination were slowed 2- to 8-fold, indicating its role in regulating olfactory response kinetics and termination. This phenotype persisted in OMP(-/-) ORNs even in low external Ca(2+) solution chosen to prevent Cl(-) channel activation, suggesting OMP acts upstream of Cl(-) channel activation. Furthermore, the response kinetics in cilia are virtually indistinguishable between OMP(-/-) and wild-type ORNs when intracellular cAMP level was elevated by the phospho-diesterase inhibitor, IBMX, suggesting OMP acts upstream of cAMP production. Together, our results suggest a role for OMP in regulating the kinetics and termination of olfactory responses, implicating a novel mechanism for fast and robust response termination to ensure the temporal resolution of the odour stimulus. These observations also help explain the deficits in odour detection threshold and odour quality discrimination seen in the OMP(-/-) mice.
Figures
Similar articles
-
The cyclic AMP signaling pathway in the rodent main olfactory system.Cell Tissue Res. 2021 Jan;383(1):429-443. doi: 10.1007/s00441-020-03391-7. Epub 2021 Jan 15. Cell Tissue Res. 2021. PMID: 33447881 Review.
-
Olfactory marker protein directly buffers cAMP to avoid depolarization-induced silencing of olfactory receptor neurons.Nat Commun. 2020 May 4;11(1):2188. doi: 10.1038/s41467-020-15917-2. Nat Commun. 2020. PMID: 32366818 Free PMC article.
-
The Odorant Receptor-Dependent Role of Olfactory Marker Protein in Olfactory Receptor Neurons.J Neurosci. 2016 Mar 9;36(10):2995-3006. doi: 10.1523/JNEUROSCI.4209-15.2016. J Neurosci. 2016. PMID: 26961953 Free PMC article.
-
Oscillatory current responses of olfactory receptor neurons to odorants and computer simulation based on a cyclic AMP transduction model.Chem Senses. 2002 Nov;27(9):789-801. doi: 10.1093/chemse/27.9.789. Chem Senses. 2002. PMID: 12438204
-
The functional relevance of olfactory marker protein in the vertebrate olfactory system: a never-ending story.Cell Tissue Res. 2021 Jan;383(1):409-427. doi: 10.1007/s00441-020-03349-9. Epub 2021 Jan 15. Cell Tissue Res. 2021. PMID: 33447880 Free PMC article. Review.
Cited by
-
Heterotrimeric G protein subunit Gγ13 is critical to olfaction.J Neurosci. 2013 May 1;33(18):7975-84. doi: 10.1523/JNEUROSCI.5563-12.2013. J Neurosci. 2013. PMID: 23637188 Free PMC article.
-
Olfactory organ of Octopus vulgaris: morphology, plasticity, turnover and sensory characterization.Biol Open. 2016 May 15;5(5):611-9. doi: 10.1242/bio.017764. Biol Open. 2016. PMID: 27069253 Free PMC article.
-
Postnatal experience modulates functional properties of mouse olfactory sensory neurons.Eur J Neurosci. 2012 Aug;36(4):2452-60. doi: 10.1111/j.1460-9568.2012.08170.x. Epub 2012 Jun 15. Eur J Neurosci. 2012. PMID: 22703547 Free PMC article.
-
The cyclic AMP signaling pathway in the rodent main olfactory system.Cell Tissue Res. 2021 Jan;383(1):429-443. doi: 10.1007/s00441-020-03391-7. Epub 2021 Jan 15. Cell Tissue Res. 2021. PMID: 33447881 Review.
-
Olfactory marker protein interacts with adenosine nucleotide derivatives.Biochem Biophys Rep. 2021 Jan 12;25:100887. doi: 10.1016/j.bbrep.2020.100887. eCollection 2021 Mar. Biochem Biophys Rep. 2021. PMID: 33490644 Free PMC article.
References
-
- Biffo S, Grillo M, Margolis FL. Cellular localization of carnosine-like and anserine-like immunoreactivities in rodent and avian central nervous system. Neuroscience. 1990;35:637–651. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous