

A novel method for poly(A) fractionation reveals a large population of mRNAs with a short poly(A) tail in mammalian cells
- PMID: 17933768
- PMCID: PMC2095794
- DOI: 10.1093/nar/gkm830
A novel method for poly(A) fractionation reveals a large population of mRNAs with a short poly(A) tail in mammalian cells
Abstract
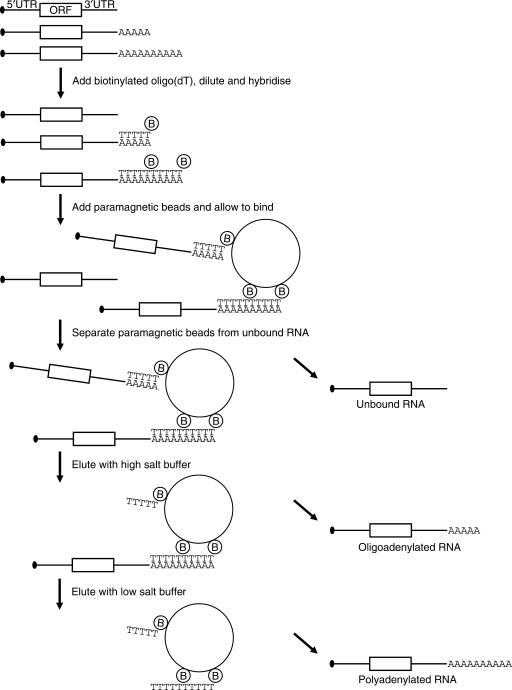
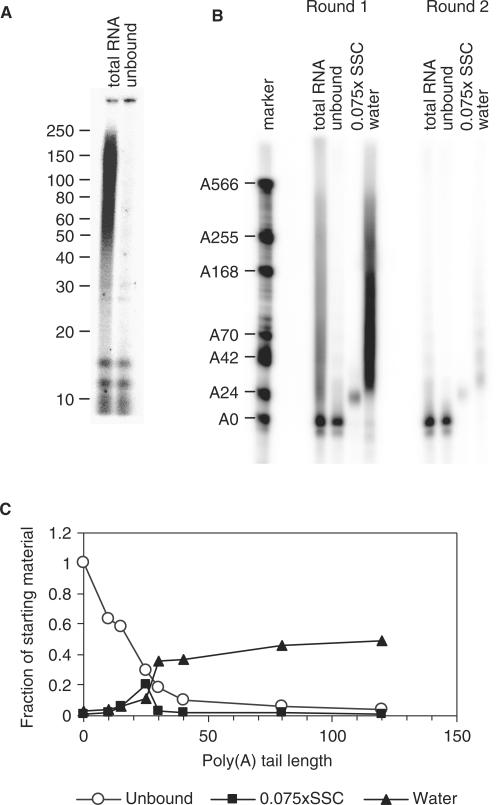
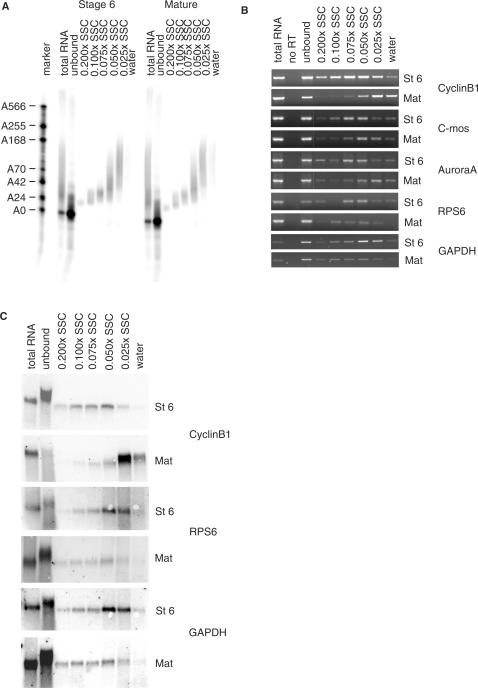
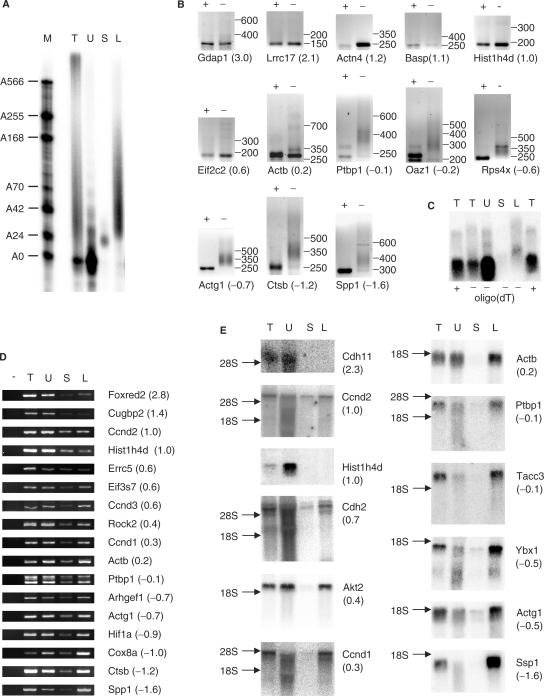
The length of the poly(A) tail of an mRNA plays an important role in translational efficiency, mRNA stability and mRNA degradation. Regulated polyadenylation and deadenylation of specific mRNAs is involved in oogenesis, embryonic development, spermatogenesis, cell cycle progression and synaptic plasticity. Here we report a new technique to analyse the length of poly(A) tails and to separate a mixed population of mRNAs into fractions dependent on the length of their poly(A) tails. The method can be performed on crude lysate or total RNA, is fast, highly reproducible and minor changes in poly(A) tail length distribution are easily detected. We validated the method by analysing mRNAs known to undergo cytoplasmic polyadenylation during Xenopus laevis oocyte maturation. We then separated RNA from NIH3T3 cells into two fractions with short and long poly(A) tails and compared them by microarray analysis. In combination with the validation experiments, the results indicate that approximately 25% of the expressed genes have a poly(A) tail of less than 30 residues in a significant percentage of their transcripts.
Figures
Similar articles
-
Control of poly(A)-tail length and translation in vertebrate oocytes and early embryos.Dev Cell. 2024 Apr 22;59(8):1058-1074.e11. doi: 10.1016/j.devcel.2024.02.007. Epub 2024 Mar 8. Dev Cell. 2024. PMID: 38460509
-
Identification of post-transcriptionally regulated Xenopus tropicalis maternal mRNAs by microarray.Nucleic Acids Res. 2006 Feb 7;34(3):986-95. doi: 10.1093/nar/gkj492. Print 2006. Nucleic Acids Res. 2006. PMID: 16464828 Free PMC article.
-
The molecular basis of coupling between poly(A)-tail length and translational efficiency.Elife. 2021 Jul 2;10:e66493. doi: 10.7554/eLife.66493. Elife. 2021. PMID: 34213414 Free PMC article.
-
Cytoplasmic mRNA polyadenylation and translation assays.Methods Mol Biol. 2006;322:183-98. doi: 10.1007/978-1-59745-000-3_13. Methods Mol Biol. 2006. PMID: 16739724 Review.
-
To polyadenylate or to deadenylate: that is the question.Cell Cycle. 2010 Nov 15;9(22):4437-49. doi: 10.4161/cc.9.22.13887. Epub 2010 Nov 15. Cell Cycle. 2010. PMID: 21084869 Free PMC article. Review.
Cited by
-
Age-related alterations in fertilization-induced Ca2+ oscillations depend on the genetic background of mouse oocytes†.Biol Reprod. 2020 Oct 29;103(5):986-999. doi: 10.1093/biolre/ioaa139. Biol Reprod. 2020. PMID: 32761132 Free PMC article.
-
Specificity factors in cytoplasmic polyadenylation.Wiley Interdiscip Rev RNA. 2013 Jul-Aug;4(4):437-61. doi: 10.1002/wrna.1171. Wiley Interdiscip Rev RNA. 2013. PMID: 23776146 Free PMC article. Review.
-
Changes in poly(A) tail length dynamics from the loss of the circadian deadenylase Nocturnin.Sci Rep. 2015 Nov 20;5:17059. doi: 10.1038/srep17059. Sci Rep. 2015. PMID: 26586468 Free PMC article.
-
The non-canonical poly(A) polymerase FAM46C acts as an onco-suppressor in multiple myeloma.Nat Commun. 2017 Sep 20;8(1):619. doi: 10.1038/s41467-017-00578-5. Nat Commun. 2017. PMID: 28931820 Free PMC article.
-
Enhanced stability and polyadenylation of select mRNAs support rapid thermogenesis in the brown fat of a hibernator.Elife. 2015 Jan 27;4:e04517. doi: 10.7554/eLife.04517. Elife. 2015. PMID: 25626169 Free PMC article.
References
-
- Piccioni F, Zappavigna V, Verrotti AC. Translational regulation during oogenesis and early development: the cap-poly(A) tail relationship. Crit. Rev. Biol. 2005;328:863–881. - PubMed
-
- Kuhn U, Wahle E. Structure and function of poly(A) binding proteins. Biochim. Biophys. Acta. 2004;1678:67–84. - PubMed
-
- Meyer S, Temme C, Wahle E. Messenger RNA turnover in eukaryotes: pathways and enzymes. Crit. Rev. Biochem. Mol. Biol. 2004;39:197–216. - PubMed
-
- Wilusz CJ, Wormington M, Peltz SW. The cap-to-tail guide to mRNA turnover. Nat. Rev. Mol. Cell Biol. 2001;2:237–246. - PubMed
-
- Giraldez AJ, Mishima Y, Rihel J, Grocock RJ, Van DS, Inoue K, Enright AJ, Schier AF. Zebrafish MiR-430 promotes deadenylation and clearance of maternal mRNAs. Science. 2006;312:75–79. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
