

Role of Septin cytoskeleton in spine morphogenesis and dendrite development in neurons
- PMID: 17935993
- PMCID: PMC2194646
- DOI: 10.1016/j.cub.2007.09.039
Role of Septin cytoskeleton in spine morphogenesis and dendrite development in neurons
Abstract
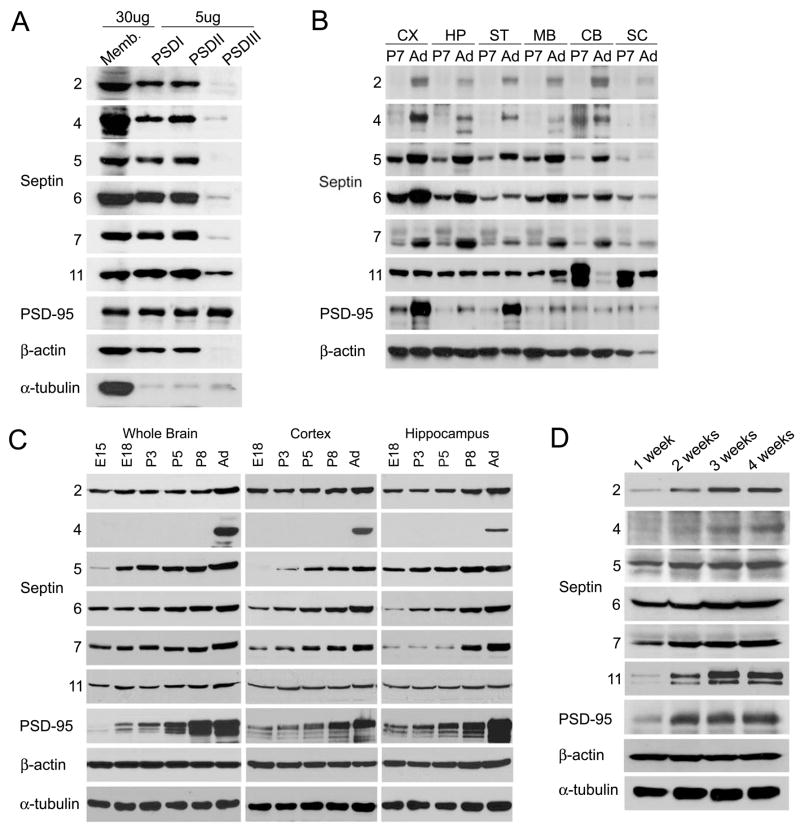
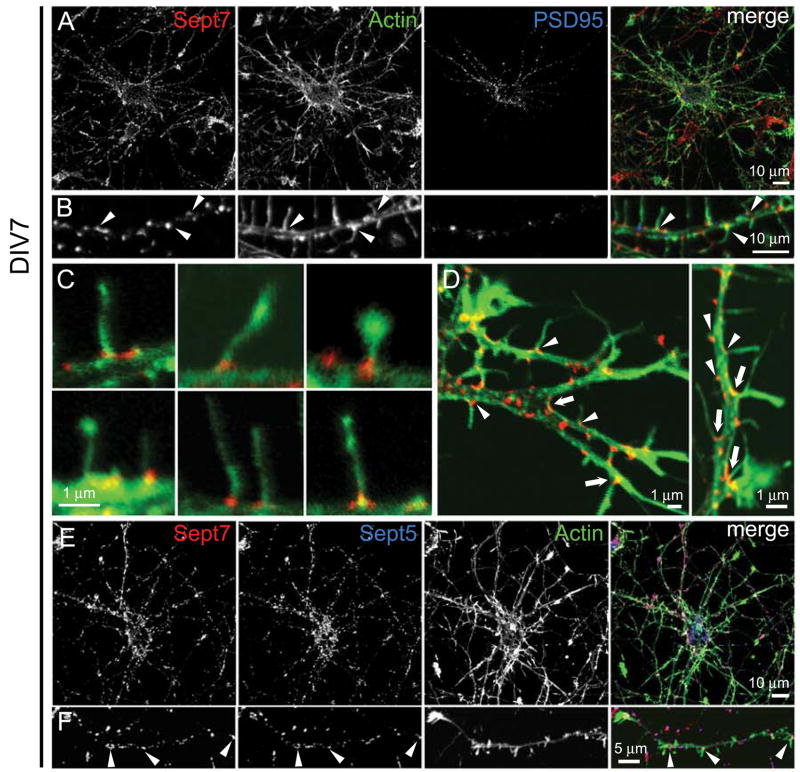
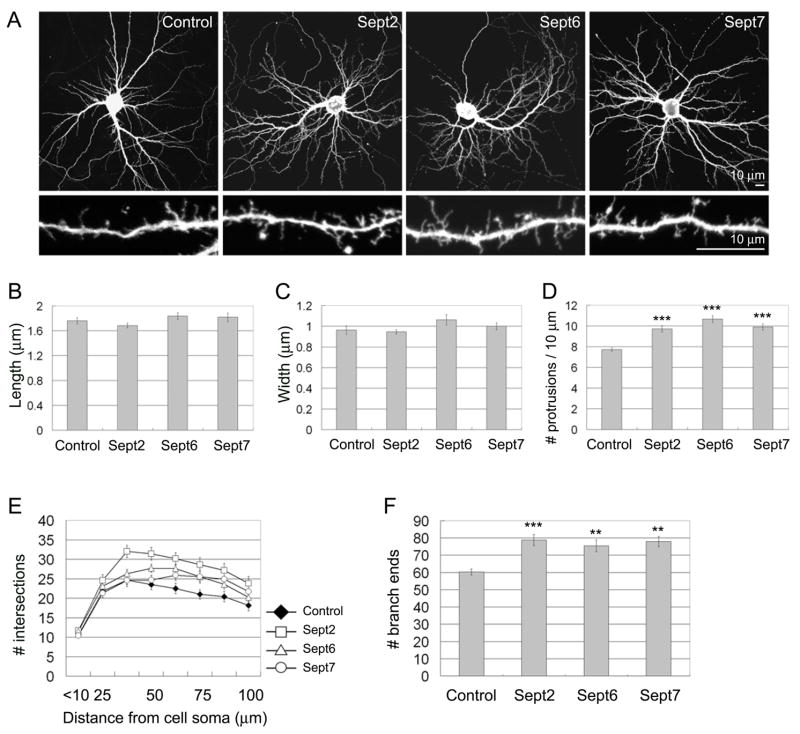
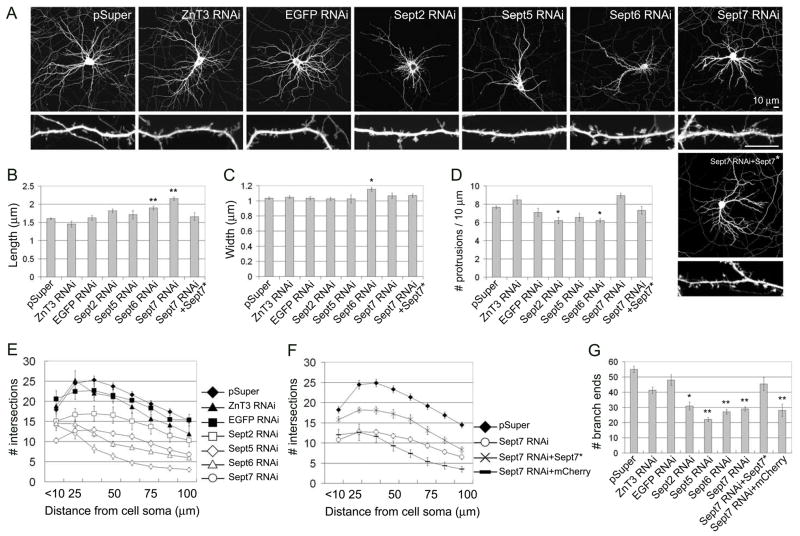
Septins are GTP-binding proteins that polymerize into heteromeric filaments and form microscopic bundles or ring structures in vitro and in vivo. Because of these properties and their ability to associate with membrane, F-actin, and microtubules, septins have been generally regarded as cytoskeletal components [1, 2]. Septins are known to play roles in cytokinesis, in membrane trafficking, and as structural scaffolds; however, their function in neurons is poorly understood. Many members of the septin family, including Septin 7 (Sept7), were found by mass-spectrometry analysis of postsynaptic density (PSD) fractions of the brain [3, 4], suggesting a possible postsynaptic function of septins in neurons. We report that Sept7 is localized at the base of dendritic protrusions and at dendritic branch points in cultured hippocampal neurons--a distribution reminiscent of septin localization in the bud neck of budding yeast. Overexpression of Sept7 increased dendrite branching and the density of dendritic protrusions, whereas RNA interference (RNAi)-mediated knockdown of Sept7 led to reduced dendrite arborization and a greater proportion of immature protrusions. These data suggest that Sept7 is critical for spine morphogenesis and dendrite development during neuronal maturation.
Figures
Comment in
-
Septins: cellular and functional barriers of neuronal activity.Curr Biol. 2007 Nov 20;17(22):R961-3. doi: 10.1016/j.cub.2007.10.001. Curr Biol. 2007. PMID: 18029249 Review.
References
-
- Kinoshita M, Field CM, Coughlin ML, Straight AF, Mitchison TJ. Self- and actin-templated assembly of Mammalian septins. Dev Cell. 2002;3:791–802. - PubMed
-
- Kinoshita M. Diversity of septin scaffolds. Curr Opin Cell Biol. 2006;18:54–60. - PubMed
-
- Peng J, Kim MJ, Cheng D, Duong DM, Gygi SP, Sheng M. Semiquantitative proteomic analysis of rat forebrain postsynaptic density fractions by mass spectrometry. J Biol Chem. 2004;279:21003–21011. - PubMed
-
- Collins MO, Husi H, Yu L, Brandon JM, Anderson CN, Blackstock WP, Choudhary JS, Grant SG. Molecular characterization and comparison of the components and multiprotein complexes in the postsynaptic proteome. J Neurochem 2005 - PubMed
-
- Hall PA, Jung K, Hillan KJ, Russell SE. Expression profiling the human septin gene family. J Pathol. 2005;206:269–278. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
