

C. elegans Enabled exhibits novel interactions with N-WASP, Abl, and cell-cell junctions
- PMID: 17935994
- PMCID: PMC2045632
- DOI: 10.1016/j.cub.2007.09.033
C. elegans Enabled exhibits novel interactions with N-WASP, Abl, and cell-cell junctions
Abstract
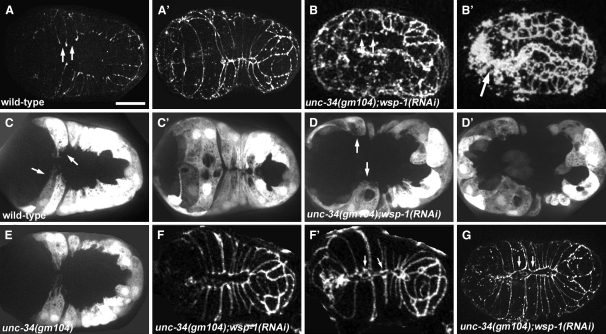
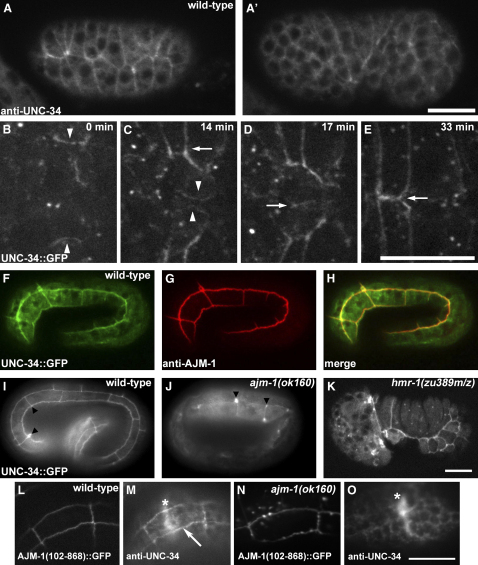
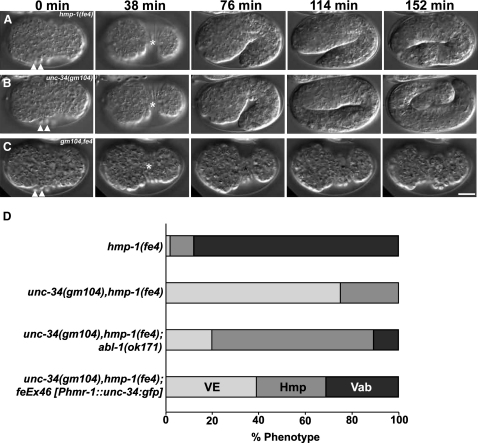
Ena/VASP proteins are associated with cell-cell junctions in cultured mammalian cells [1] and Drosophila epithelia [2, 3], but they have only been extensively studied at the leading edges of migratory fibroblasts, where they modulate the protrusion of the leading edge [4]. They act by regulating actin-filament geometry, antagonizing the effects of actin-capping protein [5]. Embryos lacking the C. elegans Ena/VASP, UNC-34, display subtle defects in the leading edges of migrating epidermal cells but undergo normal epidermal morphogenesis. In contrast, embryos lacking both UNC-34 and the C. elegans N-WASP homolog have severe defects in epidermal morphogenesis, suggesting that they have parallel roles in coordinating cell behavior. GFP-tagged UNC-34 localizes to the leading edges of migrating epidermal cells, becoming redistributed to new junctions that form during epidermal-sheet sealing. Consistent with this, UNC-34 contributes to the formation of cadherin-based junctions. The junctional localization of UNC-34 is independent of proteins involved in Ena/VASP localization in other experimental systems; instead, junctional distribution depends upon the junctional protein AJM-1. We also show that Abelson tyrosine kinase, a major regulator of Enabled in Drosophila, is not required for UNC-34/Ena function in epithelia. Instead, our data suggest that Abelson kinase acts in parallel to UNC-34/Ena, antagonizing its function.
Figures
Similar articles
-
The role of C. elegans Ena/VASP homolog UNC-34 in neuronal polarity and motility.Dev Biol. 2010 Aug 1;344(1):94-106. doi: 10.1016/j.ydbio.2010.04.025. Epub 2010 May 7. Dev Biol. 2010. PMID: 20452341 Free PMC article.
-
Caenorhabditis elegans WASP and Ena/VASP proteins play compensatory roles in morphogenesis and neuronal cell migration.Genetics. 2004 Jul;167(3):1165-76. doi: 10.1534/genetics.103.025676. Genetics. 2004. PMID: 15280232 Free PMC article.
-
Cooperative regulation of AJM-1 controls junctional integrity in Caenorhabditis elegans epithelia.Nat Cell Biol. 2001 Nov;3(11):983-91. doi: 10.1038/ncb1101-983. Nat Cell Biol. 2001. PMID: 11715019
-
The cytoskeleton and epidermal morphogenesis in C. elegans.Exp Cell Res. 2004 Nov 15;301(1):84-90. doi: 10.1016/j.yexcr.2004.08.017. Exp Cell Res. 2004. PMID: 15501449 Review.
-
The cadherin superfamily.WormBook. 2005 Dec 29:1-9. doi: 10.1895/wormbook.1.50.1. WormBook. 2005. PMID: 18050421 Free PMC article. Review.
Cited by
-
Cadherins and their partners in the nematode worm Caenorhabditis elegans.Prog Mol Biol Transl Sci. 2013;116:239-62. doi: 10.1016/B978-0-12-394311-8.00011-X. Prog Mol Biol Transl Sci. 2013. PMID: 23481198 Free PMC article. Review.
-
Cadherin complexity: recent insights into cadherin superfamily function in C. elegans.Curr Opin Cell Biol. 2012 Oct;24(5):695-701. doi: 10.1016/j.ceb.2012.06.008. Epub 2012 Jul 19. Curr Opin Cell Biol. 2012. PMID: 22819515 Free PMC article. Review.
-
Metastasis: tumor cells becoming MENAcing.Trends Cell Biol. 2011 Feb;21(2):81-90. doi: 10.1016/j.tcb.2010.10.001. Epub 2010 Nov 9. Trends Cell Biol. 2011. PMID: 21071226 Free PMC article. Review.
-
The role of C. elegans Ena/VASP homolog UNC-34 in neuronal polarity and motility.Dev Biol. 2010 Aug 1;344(1):94-106. doi: 10.1016/j.ydbio.2010.04.025. Epub 2010 May 7. Dev Biol. 2010. PMID: 20452341 Free PMC article.
-
Abl kinase inhibits the engulfment of apoptotic [corrected] cells in Caenorhabditis elegans.PLoS Biol. 2009 Apr 28;7(4):e99. doi: 10.1371/journal.pbio.1000099. PLoS Biol. 2009. PMID: 19402756 Free PMC article.
References
-
- Vasioukhin V., Bauer C., Yin M., Fuchs E. Directed actin polymerization is the driving force for epithelial cell-cell adhesion. Cell. 2000;100:209–219. - PubMed
-
- Baum B., Perrimon N. Spatial control of the actin cytoskeleton in Drosophila epithelial cells. Nat. Cell Biol. 2001;3:883–890. - PubMed
-
- Bear J.E., Loureiro J.J., Libova I., Fassler R., Wehland J., Gertler F.B. Negative regulation of fibroblast motility by Ena/VASP proteins. Cell. 2000;101:717–728. - PubMed
-
- Bear J.E., Svitkina T.M., Krause M., Schafer D.A., Loureiro J.J., Strasser G.A., Maly I.V., Chaga O.Y., Cooper J.A., Borisy G.G., Gertler F.B. Antagonism between Ena/VASP proteins and actin filament capping regulates fibroblast motility. Cell. 2002;109:509–521. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
